
Данная информация предназначена для специалистов в области здравоохранения и фармацевтики. Пациенты не должны использовать эту информацию в качестве медицинских советов или рекомендаций.
Механизмы генного многообразия в геномах
М.А. Гайдес, к.м.н.
Заведующий лабораторией кардио-респрираторного тестирования, Госпиталь им. Хаима
Шиба, Тель Ашомер, Израиль,
Address: 20 Hamelech Rachavam str, 3, Ashdod, 7748019, Israel.
gaidesmark@gmail.com
Известно, что в начальных фазах развития Земли (около
4.5 миллиардов лет назад) на ней вообще не было ничего живого. Затем на
протяжении примерно 3-х миллиардов лет на Земле живые организмы были
представлены только вирусами и прокариотами, имеюших одно- или малогенные геномы.
И только около 500-600 миллионов лет назад появились многоклеточные растения и
животные, имеющие многогенные геномы. Также известно, что чем сложнее построена
особь, тем больше генов содержит её геном. Следовательно, биологическая эволюция
нарастала только за счёт прироста числа генов в геномах.
Но также известно, что ни одна биологическая особь не может сама «выращивать»
свои гены. Следовательно, существует проблема объяснения, каким образом
происходил прирост генов в геномах биологических особей.
В данной статье предлагается механизм наращивания числа генов в геномах за
счёт механизма генной мультипликации. Этот механизм включает в себя два
отдельных механизма – механизм вирусного заражения и механизм полового
скрещивания. Оба механизма предназначены для наращивания числа вирусных генов в
геномах особей всех биологических видов.
Биологическая эволюция всегда происходит только путем усложнения генома видов
– прибавления новых генов к уже имеющимся. Это значит, что каждый новый вид
происходит из старого только путем появления новых генов в геноме старого вида.
Вся эволюция биосистем за все время существования Земли осуществлялась только за
счет прибавления новых генов в геномы родительских видов. При этом возникает
вопрос: – откуда берутся в геномах живых организмов дополнительные гены?
Наращивание генов в геноме.
Цель любого живого объекта – размножение, которое является действием,
осуществляемым с помощью неких средств (инструментов). Этими средствами являются
биологические конструкции (мембраны, жгутики, энзимы, внутриклеточные органеллы,
внеклеточные образования и прочее), которые синтезируются самими живыми
объектами. Каждая биоконструкция построена либо из одной или множества молекул
белка, либо при обязательном их участии. Синтез каждой молекулы белка
контролируется соответствующим геном по принципу – «один ген – одна молекула
белка». Возможно механизм сплайсинг кода позволяет некому числу генов управлять
синтезом большего числа белков. Но всегда для синтеза любых молекул белка
необходимы некие гены. Ни одной молекулы белка не появится без соответствующих
генов. Следовательно, сколько типов молекул белка требуют те или иные
биологические конструкции, столько генов должно находиться в геноме.
Генόм является той основой, которая определяет все свойства живых систем,
потому что в нем «записана» технология строительства биологических элементов и
их сборки в биологические конструкции. Только он определяет синтез этих
конструкций, которые определяют все свойства биосистем. Следовательно, синтез
любых биологических конструкций зависит только от наличия соответствующих генов.
Чем больше генов, тем больше можно построить биологических конструкций, тем
больше устойчивость биосистемы во внешней среде, тем лучше она выполняет свою
цель – размножение. Поэтому рассмотрение теорий эволюции невозможно без
рассмотрения роли генома для развития и функционирования биосистем.
Но в биологии существуют две проблемы – проблема появления первого гена на
Земле и проблема наращивания числа генов в геноме.
Проблема происхождения первого гена решается с помощью гипотез о минеральном
происхождении генов – гипотез о существовании в далёком прошлом мира РНК.[5,
16, 19, 22] Эта проблема сама состоит из двух подпроблем: проблемы
происхождения рибонуклеотидов, которые не могли быть синтзированы в условиях
первобытной Земли, и проблемы матрицы для синтеза РНК из рибонуклеотидов.
Проблема происхождения рибонуклеотидов решается с помощью гипотез о том, что
первые полинуклеотиды могли быть синтезированы не из рибонуклеиновых кислот, из
которых синтезируются молекулы РНК и ДНК, а из неких других мононуклеотидов (ПНК
– peptide nucleic acide, [24] ТНК – threose
nucleic
acide . [26] и ГНК –
Glycerol
nucleic acide ,
[1, 27]), которые затем эволюцией были заменены на рибонуклеотиды. Также
есть работы, в которых показано, что рибонуклеотиды могли быть синтезированы не
из рибозы и азотистых оснований, а из более простых органических молекул, таких
как формальдегид. Возможно, в ходе химической эволюции между простейшими
органическими веществами и РНК вообще не было промежуточного этапа накопления
сахаров и азотистых оснований.[22]
Проблема матрицы для синтеза РНК из рибонуклеотидов решается с помощью
гипотез о том, что первичной матрицей, на которой могли быть синтезированы
полинуклеотиды (РНК и ДНК), могли быть некие кристаллы [9, 34, 39],
глины [4] и другие. Эти гипотезы ещё не доказаны, но в любом случае у
нас есть хотя бы некоторое представление о том, каким образом мог появиться
первый ген.
Вторая проблема (происхождение многогенности) решается намного хуже. Мы не
знаем, откуда в геномах вообще взялось множество генов. На заре биологической
эволюции первые прото-клетки (вирусы) имели считанное число генов. А спустя
миллиард лет эволюции из прото-клеток появились первые клетки, в геномах которых
было уже около 1000 генов. Если клетки произошли от малогенных вирусов, то каким
образом произошло увеличение числа генов в геноме? Существует множество гипотез,
пытающихся объяснить эволюцию на основе генетики и теории Дарвина.
В начале ХХ века Рональд Фишер, Джон Холдейн-младший и Сьюэл Райт
сформулировали синтетическую теорию эволюции (СТЭ) в её нынешнем виде. Толчок к
развитию СТЭ дала гипотеза о рецессивности новых генов, которая предполагала,
что в каждой воспроизводящейся группе организмов во время созревания гамет в
результате ошибок при репликации ДНК постоянно возникают мутации – новые
варианты генов. Плейотропия и полимерия отражают взаимодействие генов, благодаря
которому внешнее проявление каждого гена зависит от его генетического окружения.
В результате рекомбинация, порождая все новые генные сочетания, проявляется в
фенотипе особи-носителя. Далее естественный отбор уничтожает те сочетания генов,
которые затрудняют жизнь и размножение организмов в данной среде, и сохраняет
нейтральные и выгодные сочетания, которые подвергаются дальнейшему размножению,
рекомбинации и тестированию отбором.[ 13, 35, 37]
В вопросе об источнике генетического разнообразия СТЭ признает главную роль
за мутацией и рекомбинацией генов. Но чтобы гены мутировали, они изначально
должны быть в геноме во множестве! Рекомбинация изначально предполагает
многогенность – чтобы фрагменты генома могли мигрировать из клетки в клетку, они
сначала должны быть в геноме. Но если уже есть генное многообразие и процесс
рекомбинации генов, то нет необходимости в привлечении гипотезы мутагенеза!
Кроме того, СТЭ предполагает медленную поэтапную эволюцию с постепенным
изменением генома и множеством переходных форм, а в реальности мы наблюдаем
скачкообразность эволюции видов. Если следовать современным утверждениям теории
эволюции, то окаменелые останки:
- должны иметь огромное число так называемых переходных форм жизни,
- из-за наличия переходных форм организмы должны развиваться очень медленно
и поэтапно, от примитивных форм ко все более сложным формам жизни,
- по мере развития и появления новых клеток, должны появляться новые виды
жизни,
- останки которых непременно должны быть среди окаменелостей.
Ни одно из этих утверждений СТЭ в ископаемых останках не нашло никакого
подтверждения фактами. Самый древний из найденных окаменелых останков мечехвоста
возрастом 450 миллионов лет, относящийся к Ордовикскому периоду, доказывает, что
почти за полмиллиарда лет это существо не претерпело ни единого анатомического
или биологического изменения и существует и поныне в точности таким, каким оно
было сотни миллионов лет назад, с момента своего появления на Земле. Однако в
периоды ранее 500 милиионов лет назад мечехвост не обнаруживается. Это означает,
что первый же мечехвост появился сразу во всем своем совершенстве и у всех
потомков этого архаичного мечехвоста на протяжении сотен миллионов лет был один
и тот же синтез одних и тех же белков в одних и в тех же биохимических циклах,
управляемые совершенно одинаковыми наборами генов. Как мечехвосту на протяжении
полумиллиарда лет удалось избежать мутаций?
Все эти свидетельства дали основание утверждать, что эволюционная теория
Чарлза Дарвина неверна и все это свидетельствует о том, что формы жизни были
сотворены Всевышним Творцом сразу все и в самых совершенных формах, которые
дошли до наших дней неизменными, без следа эволюционирования.
Но эволюция существует и управляема законом Дарвина. Это видно из тех же
данных палеонтологии, которые говорят о том, что весь спектр сегодняшних
организмов появился всё же не сразу, а поэтапно – сначала более простые, а затем
более сложные. Это противоречит утверждению о единоразовом сотворении Всевышним
Творцом сразу всех видов.
Но при этом каждый вид появлялся сразу адаптированным под те внешние условия,
которые были в момент его появления, в полном соответствии с законом Дарвина. И
хотя каждый вид появлялся практически в завершенном виде, без переходных форм
(скачкообразно), каждый вид появлялся поэтапно, каждый в свое время.
Чтобы объяснить скачкообразность эволюции в 1901 году Хуго де Фризом была
предложена теория сальтационизма. Видообразование происходит очень быстро, в
течение нескольких поколений. Процесс связан с появлением новых особей, резко
отличающихся и репродуктивно изолированных от представителей родительского вида.
Эта теория основана на понятиях системных мутаций и макромутаций как особого
рода наследственных изменениях, затрагивающих всю систему генотипа («reaction
system») и несводимых к изменениям отдельных локусов.[17]
Однако теории сальтационизма и им подобные не спасают положения, потому что
они все также основаны на предположении о мутациях некого множества генов в
геноме, которые уже должны быть прежде, чем дочерний вид отпочкуется от
материнского. Кроме того, эта теория не справляется с проблемой репродуктивной
изоляции нового вида.
Даже если предположить, что изначально существует некий многогенный вид и
произошла некая «удачная» мутация генов и даже родилась особь некого нового
биологического вида, то эволюция данного нового вида будет обречена на
прекращение, потому что данная особь не найдет себе полового партнёра для
продолжения рода. Даже в самом удачном случае мутагенеза вновь появившийся
мутант обречен на видовую изоляцию и размножиться не сможет, потому что не будет
одновидового полового партнёра. А особи разных видов не скрещиваются – это
известный факт. Преодолеть репродуктивный барьер невозможно потому, что слишком
низка вероятность одновременной одинаковой мутации у двух различных особей,
чтобы появились две новые одинаковые особи нового вида, да ещё разного пола,
чтобы дать потомство.
Но даже и в этом слишком невероятном случае у этих особей нет шансов
образовать новый вид, потому что у них очень мало шансов выжить вообще, потому
что этому препятствует их малочисленность. Сегодня мы знаем вымирающие виды,
насчитывающие несколько сот особей и говорим, что они обречены на вымирание в
силу тех или иных причин, о которых здесь говорить не будем. Что же говорить о
двух особях?
Теория Дарвина и современное её продолжение (синергетика) предполагают
самоэволюцию живых систем.[33, 40] Эти теории самоадаптации
утверждают, что внешняя среда диктует условия существования живых систем, а
живые системы сами адаптируются под них. Чтобы выжить, эти системы должны
перестроить структуры своего организма таким образом, чтобы их функции были
подходящими (адаптированными) для этих условий. Если удалось это сделать,
организм выживет, если нет, то вымрет.
Однако в этих теориях самоадаптации демонстрируется полное непонимание теории
адаптации Дарвина. В них совершенно не учитывается, что новый орган, будь то
хвост, крылья, рога или когти, или даже его изменение (короткие когти вместо
длинных), могут появиться только в том случае, если в молекуле ДНК (в геноме)
уже будут присутствовать (!) соответствующие гены, которые смогут контролировать
синтез тех белков, которые построят новый орган или изменят уже существующий. В
основе синергетики лежит неверная идея САМО-усложнения, которая заключается в
том, что живые системы якобы сами вырабатывают свои приспособления к новым
условиям: мол, если мне нужны когти, то я их выращу. Но чтобы вырастить когти,
для этого нужно иметь соответствующие гены.
Концепция САМО-усложнения (синергетика) неверна именно потому, что не
учитывает роль генного механизма для синтеза адаптивных биологических
конструкций. Да, любая биологическая конструкция является адаптивной и
появляется в нем только как следствие биологической необходимости (как
облегчающая размножение в данных условиях). Но она может возникнуть только в том
случае, если для этого уже есть соответствующие гены. Если таких генов нет, то
никакая конструкция возникнуть не может, несмотря на любую необходимость в ней,
потому что сама клетка вырастить в своей молекуле ДНК по собственной инициативе
нужные гены не может.
Основной проблемой синергетики является именно неучет роли генов.
Естественно, во времена Дарвина никто ничего не знал ни о каких генах. Дарвин
наблюдал изменения в различных живых организмах, подходящие для тех условий
внешней среды, в которых они проживали и описал то, что видел. И у него понятия
не было ни о генах, ни о ДНК и ни о чём прочем подобном. А видел он только то,
что в определённых условиях выживают только адаптированные и именно к данным
условиям виды. Отсюда и был сделан вывод об адаптации живых организмов к
условиям окружающей среды. Это был наблюдаемый факт, но его интерпретация
неверна.
Почему-то ни Дарвин и никто другой не обратил внимание на то, что в этих же
условиях проживали и другие виды и не у всех были одинаковые средства адаптации.
Если бы только условия внешней среды диктовали бы форму и функции проживающих в
ней особей, то все виды без исключения должны были бы иметь одинаковые средства
адаптации (одинаковую форму), потому что они все проживают в одинаковых
условиях. Дарвин сделал неверный вывод о причинах этих адаптаций – он считал,
что сами организмы «выращивают» биологические конструкции для адаптации, «кто во
что горазд», и это объясняет различие в строении разных видов.
Однако это не так. Сами организмы не могут выращивать никаких биологических
конструкций, если у них нет для этого соответствующих генов. А «выращивать»
нужные гены они не могут. Верно, условия могут вызвать изменения в живых
организмах. Однако это может быть только в тех случаях, если в их геноме уже
есть готовые, но «дремлющие» генные наборы, способные синтезировать адекватные
внешним условиям биологические конструкции. В этом случае условия (триггер)
внешней среды могут «пробудить» эти генные наборы и те начнут управлять синтезом
соответствующих биологических конструкций, которые начнут функционировать и
адаптировать организм к этим условиям. И внешне это будет выглядеть так, как
будто сам организм адаптировался под внешние условия, вырастив новый орган и
создав новый вид. Причем это морфологическое изменение будет происходить скачком
– не было некого органа и вдруг он появился. Именно эта скачкообразность и
породила теории сальтационизма. Наличие такого скачка – это наблюдаемый факт, но
эти теории не смогли объяснить причины этого факта.
Если следовать утверждениям синергетики и присовокупить к ним генетику, то
получается странная и неестественная картина. Если клетка чувствует, что не
может противостоять внешней среде, то она неким образом передает сигнал своему
генетическому аппарату, что нужны такие-то и такие-то дополнительные гены,
кодирующие дополнительные белки. Эти гены «вырастают» на молекуле ДНК и начинают
регулировать постройку необходимых биологических конструкций. Таким образом
клетка адаптируется к новым условиям. Но это невозможно в силу ряда причин.
Во-первых, центральная догма молекулярной биологии, обобщающее и наблюдаeмоe
в природе правило реализации генетической информации, говорит об обратном –
информация передается от нуклеиновых кислот (от ДНК) к белку, но не в обратном
направлении. Правило было сформулировано Френсисом Криком ещё в 1958 году[7]
и приведено в соответствие с накопившимися к 1970 году данными.[8]
Переход генетической информации от ДНК к РНК и от РНК к белку является
универсальным для всех без исключения клеточных организмов и лежит в основе
биосинтеза макромолекул. Репликации генома соответствует информационный переход
ДНК→ДНК. В природе встречаются также переходы РНК→РНК, ДНК→РНК (трансляция) и
РНК→ДНК (у некоторых вирусов), а также изменение конформации белков,
передаваемое от молекулы к молекуле, но никогда от белка к РНК или ДНК.
Отсюда следует, что молекулы белка не могут воздействовать на молекулы ДНК и
влиять на их генный состав. Наоборот, гены ДНК определяют строение (качество)
белковых молекул, а число генов в молекуле ДНК определяет число (количество)
типов синтезируемых белков. Свет не зависит от тени, но тень зависит от света.
Есть свет – может быть тень, нет света, не может быть никакой тени. Белки
являются «тенью» ДНК.
Во-вторых, принципы общей теории систем (ОТС) [36] не позволяют
клетке самоусложняться таким образом. Эти принципы говорят о том, что ничего не
может происходить беспричинно и само собой. Для любого явления должен быть
механизм, реализующий это явление. Если принять, что клетка сама адаптируется к
новым условиям, то клетка сама должна создать:
- новые гены в молекулах РНК/ДНК, которые будут кодировать синтез
- новых белков, создающих
- новые внутриклеточные структуры (подсистемы), нужные для этих условий.
Согласно ОТС, чтобы таким путем сама клетка могла создать новые гены в ответ
на новые воздействия внешней среды, она должна:
- иметь рецепторы на эти новые воздействия, чтобы почувствовать их и
передать сигнал на ДНК о том, что нужны новые гены, хотя с этими
воздействиями она не знакома и, естественно, у неё нет этих рецепторов,
иначе эти воздействия были бы для неё не новыми,
- знать те структуры, которые ей нужны, чтобы противостоять этим
воздействиям. А поскольку таких структур у неё нет, то она должна:
a. знать, как построить эти структуры,
b. знать, какие белки для этого нужны,
c. знать, какие гены должны быть в её молекуле ДНК, чтобы
синтезировать эти белки и
d. уметь синтезировать эти гены в молекуле ДНК.
Другими словами, примитивная клетка должна слишком много знать и уметь, чтобы
самоусложниться, но таких знаний нет даже у человека. Поэтому процесс
САМО-усложнения генома принципиально невозможен и современное понимание
синергетики и теории адаптации Дарвина неверно.
Если принять все эти теории (СТЭ, сальтационизм, теорию адаптации,
синергетику) и подвести под них общую черту, то получается очень противоречивая
картина. Согласно ей на Земле сразу появилось множество различных живых
организмов, имеющих геномы, содержащие разное количество генов – малогенные и
многогенные. Этим геномам соответствовало определённое множество различных видов
живых систем. Затем геномы стали меняться, мутируя и вследствие этого появлялись
новые виды.
Эти теории предлагают два механизма изменения генов – мутагенез и
рекомбинацию (обмен генами или их частями), но они не предлагают никакого
механизма наращивания числа генов в геноме. Оба механизма (мутагенез и
рекомбинация) известны и наблюдаются в жизни, потому их и привлекли в качестве
основных механизмов изменчивости видов. Однако мутагенез может сработать только
тогда, когда геномы уже многогенные. Оценка роли этих механизмов в изменчивости
видов неверна, потому что основана на неверных предпосылках – на изначальной
многогенности геномов, чего не было в начале биологической эволюции.
Предполагается, что сначала появились одногенные прото-клетки (вирусы, если
верна теория мира РНК) и только потом появились многогенные прокариоты, которых
сменили ещё более многогенные одно- и многоклеточные эукариоты. Затем геномы ещё
больше «выросли» и из многоклеточных эукариотов появились многоклеточные
растения, животные и человек. Нам точно известно, что чем сложнее организм, тем
больше геном. Однако неизвестно, каким образом происходил прирост генов в
геномах, а эти теории не дают на это ответа.
К тому же, если уже с самого начала были многогенные геномы, то теории
мутагенеза вообще не нужны, потому что многогенность вкупе с рекомбинацией
вполне может дать все разнообразие геномов. Все эти теории неким образом
напоминают сотворение мира, описанное в Библии. Если с самого начала были
многогенные геномы, это значит, что с самого начала на Земле появилось некое
разнообразие многогенных живых организмов, подобно библейскому сотворению всех
живых (кроме человека) за пару дней.
У разных организмов разные наборы генов, из чего следует разный набор
адаптивных механизмов и многообразие видов живых систем. Одну и ту же цель
(размножение) разные организмы достигают разными путями (разные адаптивные
механизмы), в зависимости от имеющихся генных наборов. Осталось найти причину
появления этого набора генов в геноме биологических особей.
Сегодня принято, что основным механизмом появления новых видов является
мутация молекулы ДНК, дающая прирост числа генов с последующей передачей новых
генов половым путем своему потомству. Т.е., в наращивании числа генов участвует
два механизма:
- мутация и
- половая передача мутации.
Но мутация – это появление дефектов в «исправной» молекуле ДНК с последующим
её укорочением за счёт дефектных генов. Она объясняет только механизм появления
нарушений в генах и уменьшение их числа. Чтобы возникла мутация, в геноме уже
должны быть «лишние» гены, которые сами собой не «вырастают».
К этому следует добавить, что любая внутриклеточная конструкция требует для
своего строительства не одного, а определённого множества белков и,
следовательно, такого же, если не большего, множества генов. Появление только
одного гена не даст положительного биологического эффекта, потому что для
появления нового органа требуется появление множества генов (набора генов). В
силу редкости одиночной положительной мутации ещё труднее предположить такую
мультимутацию, которая дает положительный биологический эффект (одновременную
положительную мутацию множества генов). Механизм генной мутации не может
объяснить появление новых внутри- и внеклеточных конструкций, потому что мутация
– это всегда повреждение молекулы ДНК.
Если появляются дефектные (мутированные) гены в соматических (тканевых)
клетках многоклеточных, то появляются различные дефекты тканей – опухоли,
дистрофии, язвы и т.д. Если появляются такие гены в половых клетках, то
появляются различные дефекты развития особей – различные пороки сердца,
гипоплазии органов (недоразвитие тканей) и другие врожденные болезни типа
болезни Дауна, гемофилии, миопатии и прочие.
Чтобы появилась особь не дефектная, а наоборот, прогрессивная, с новыми
положительными качествами, нужно появление в организме новой биологической
конструкции, состоящей из многих белков и других полимеров. Для этого
недостаточно появления только одного, мутированного или немутированного гена.
Необходимо появление сразу множества новых, причём нормальных генов
(полноценного набора, комплекта генов), «запускающих» новые биохимические циклы,
способные синтезировать новые белки, из которых будут построены новые
конструкции, обеспечивающие положительный эффект. Эволюционный прирост числа
генов в ДНК от патологической мутации отличается тем, что эволюционный прирост
добавляет новые гены, создавая их избыточность, а патологическая мутация
уменьшает число нормально работающих генов. Однако неизвестен механизм
эволюционного прироста генов в геномах.
Постройка новых белков и всего остального возможна только тогда, когда в
молекуле ДНК готовые для этого гены уже есть. Несоответствие теорий мутагенеза
естественному отбору Дарвина в том, что ДНК заведомо должна содержать в том или
ином виде (в латентном, например, в «мусорных» генах) информацию о технологии
производства новых белков. Эта информация может быть записана только в генах. И
опять тот же вопрос: – откуда гены появляются в геноме?
В сегодняшней биологии ответа на этот вопрос нет и существует проблема
объяснения наращивания числа генов в геноме. Теория мутагенеза не проходит
потому, что она подразумевает мутацию генов в геноме, которые в нем уже должны
быть. И она не объясняет, откуда в нем появились эти гены, которые должны или не
должны мутировать. Поэтому предлагается теория генной мультипликации.
Генная мультипликация
Генная мультипликация – это процесс присоединения новых генов к молекуле ДНК
путем вирусного заражения или половой передачи вирусных генов.
На накопление избыточных генов и их включение в функцию, т.е., настройку
генных наборов законом Дарвина, было затрачено около 3-4 миллиардов лет.
Существует множество механизмов появления новых генов в геноме. Основных из них
два:
- вирусное заражение с последующей рекомбинацией и встраиванием вирусного
гена в геном клетки-хозяина и
- половое скрещивание.
Процесс вирусного заражения – это механизм размножения вирусов. При таком
заражении вирусы встраивают свой геном в геном жертвы, вирусный геном начинает
конкурировать с геномом жертвы и вырабатывать биологические конструкции вируса.
Описано несколько типов взаимодействия между вирусом и клеткой:
- лизис,
- лизогения,
- псевдолизогения и
- хроническая инфекция (Weinbauer, 2004).[29]
При лизисе происходит гибель клетки и выход вирусных частиц в окружающую
среду немедленно после латентного периода (периода формирования вирусов). При
лизогении вирусный геном в неинфекционной форме становится частью генома хозяина
или сохраняет самостоятельность и передается зараженными клетками потомству.
Вирусный геном существует в покоящейся форме (профага) до наступления лизиса,
вызываемого внешними факторами (Wilson, Mann, 1997; Fuhrman, 1999).[15, 30]
Лизогению вызывают умеренные вирусы, которые играют большую роль в
горизонтальном генетическом переносе информации – трансдукции. При
псевдолизогении вирусный геном в неактивном состоянии находится в клетке в
течение длительного периода, но не интегрируется с геномом хозяина и не способен
к репликации.
Предполагается, что псевдолизогения возникает при сильном истощении клетки.
Однако при наступлении благоприятных условий для вируса состояние
псевдолизогении прекращается, вызывая настоящую лизогению и инициируя литическую
продукцию вирионов. При хронической инфекции клетки хозяина не лизируются в
течение вирусного жизненного цикла, а живые бактерии освобождают вирусы из
клетки путем отпочковывания.[38] Но что кроется за каждым из этих
процессов?
Геном вируса Megavirus chilensis (рис. вв) состоит из почти 1.26 миллиона пар
нуклеотидов (рис. 1).
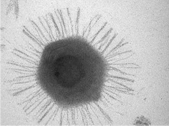
Рис. 1. Вирус Megavirus chilensis [фото с сайта http://news.rambler.ru].
На сегодняшний день он самый длинный среди известных человеку геном[11]
вируса. В геноме этого мегавируса есть 1120 генов, а в геноме его более
«мелкого» родственника ( Acanthamoeba
polyphaga
mimivirus ) – 979 генов. Откуда все эти
гены? Из мутаций? Но чтобы мутировать ген должен присутствовать в геноме!
Такое «богатство» генов объясняют тем, что мимивирус – это «...чрезвычайно
успешный генетический «карманник», большой геном которого объясняется
горизонтальным переносом генов от амеб, бактерий или других вирусов...».[11]
Существует точка зрения, объясняющая появление мимивируса из мегавируса –
редукционизм.[11] Согласно этому сценарию, доля генов, полученных в
результате горизонтального переноса, не превышает 10%. И из этого делается вывод
– этот относительно сложный геном мимивирус «унаследовал» от ещё более сложного
организма-предка! Исходя из того факта, что мегавирус и мимивирус содержат
большое количество идентичных генов был сделан вывод, что первоначально был
некий предок – клеточный организм, который путем постепенной деградации генома
(редукции) сначала превратился в мегавируса, а затем «потеряв» ещё несколько
генов, превратился в мимивируса.
Тогда почему бы не продолжить эту же логику и не предположить, что более
мелкие вирусы произошли уже из мимивирусов и так до одногенных вирусов. Применяя
эту же логику можно утверждать, что вообще все вирусы произошли от прокариотов,
прокариоты произошли от эукариотов, те от многоклеточных, а эти из растений,
животных и т.д. Т.е., все можно поставить с ног на голову в буквальном смысле –
сначала появился человек, а затем его геном стал рассыпаться и, путем
деградации, появились все остальные более простые системы биотических миров,
вплоть до вирусов, по принципу появления миллионера из разоряющегося
миллиардера. Остается только один вопрос: – откуда появился сам миллиардер?
Такая логика полна противоречий. Если «рассыпать» мегавирус на отдельные
гены, то получим всего лишь 1200 генов, в то время как число видов вирусов, а
значит и генов, составляет сотни миллионов, если не миллиарды. Их настолько
много, что даже самого богатого генома и даже суммы генов всех геномов живых
организмов не хватит, чтобы «перекрыть» все многообразие геномов вирусов. Не
проще ли предположить, что сам мегавирус неким определённым образом был собран
из 1200 отдельных вирусов?
Есть веские причины усомниться в том, что мимивирус «украл» гены у амёб,
бактерий и т.д. Мы знаем обратный процесс, когда вирус встраивает свой геном в
геномы более развитых живых систем (тех же амёб, бактерий и т.д.), но нам
незнаком обратный процесс, чтобы более развитые живые системы (амёбы и бактерии)
встраивали свой геном или его части в геном более низкоразвитых живых систем (в
вирусы).
Считается, что вирусы, как правило, всегда паразиты – фаги. Вирусы, которые
встраивают свой геном в геномы других вирусов – это вириофаги, бактерий –
бактериофаги. Но никто ещё не видел амёб или бактерий-вирусофагов.
Известен факт наличия мелких вирусов в других вирусах, которые живут в
клетках-хозяевах. Эти вирусы считаются спутниками этих других вирусов. Эти новые
вирусы считаются вредными для их так называемых принимающих вирусов, которые
становятся как бы больными. Поэтому было принято понятие virophage. Вирофаг –
это вирус, который заражает другой вирус. Исследования по морфологии, жизненного
цикла и геномов virophages обогатили эту концепцию, выделив неожиданные
особенности. Таким образом, появился класс virophages среди спутниковых вирусов
в рамках virosphere.[23]
Например, известны три вириофага:[12, 20 и другие]
- Спутник (Sputnic virophage) – паразитирует на мимивирусе амёбы
Acanthamoeba polyphaga.
- Мавирус (Mavirus) – паразитирует на Cafeteria roenbergensis жгутиконосца.
- OLV (Organic Lake Virophage), обнаружен а Антарктиде – паразитирует на
вирусах Phycodnaviridae, которые обитают в водорослях хлореллы, которые, как считается, паразитируют на других вирусах (рис. 2).
![Рис. 2. Маленькие вирусы-спутники (как считают – паразиты) внутри гигантского мимивируса. Длина масштабной линейки�— 200�нм.[2]](images/art/all20/new_pa13.jpg)
Рис. 2. Маленькие вирусы-спутники (как считают – паразиты) внутри
гигантского мимивируса. Длина масштабной линейки�— 200�нм.[2]
Однако вирус не может паразитировать на вирусе, потому что любой вирус ничего
не производит, кроме своих РНК/ДНК или молекул белка, которые при таком
«паразитировании» остаются целыми и у «паразита», и у вируса-жертвы. Может быть
вириофаги и мешают мимиврусам жить, потому что вмешиваются в порядок генов в их
геномах, но они ничего «не едят» внутри них, потому что у них нет элементов
системы пищеварения, позволяющий им «переваривать» ДНК и белки мимивирусов и
расщеплять эти молекулы на моно-нуклеотиды и аминокислоты.
Примеры взаимодействий вирусов с вирусами говорят о том, что вирусы неким
образом объединяют свои геномы, увеличивая число генов в них (рис. 3). Такое
объединение называется вирусной коньюгацией, которую можно назвать иначе –
генной мультипликацией, конечным результатом которой и появилось всё
разнообразие многогенных вирусов. Благодаря генной мультипликации сложность
генома вирусов нарастала и результатом этого было появление прокариотов –
полноценных безъядерных клеток. Дальнейшая коньюгация молекул ДНК, основанная на
том или ином механизме, привела к дальнейшему нарастанию генома и появлению всех
последующих уже ядерных организмов (эукариот), одно- и многоклеточных.
Мимивирус и мегавирус имеют около 594 белков, сходных по структуре и функции.
При этом в геноме первого закодировано 979 белков, а второго — 1120. Это говорит
о том, что и у вирусов уже стала набираться генная избыточность, когда генов
больше, чем синтезируемых под их контролем белков. Единственный механизм,
который может объяснить нарастание генома вирусов – это вирусная конъюгация,
вернее, ДНК-конъюгация.
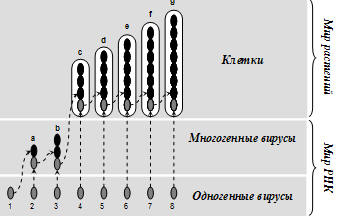
Рис. 3. Генная мультипликация. Появление и накопление генной избыточности
за счет коньюгации генов через вирусное заражение.
Каждый ген изображен в
виде черных вытянутых точек-элипсов (от 1-го до 8-го). Белые вытянутые овалы
большего размера – отдельные клетки (c, d. e, f, g). Вирус «а» родственен
одногенным вирусам «1» и «2», потому что произошел за счет коньюгации их
генов. Но он не родственен всем остальным вирусам, потому что не имеет их
генов. Вирус «b» родственен одногенным вирусам «1», «2» и «3», а также
многогенному вирусу «а», потому что его геном включает в себя их гены, но не
родственен остальным вирусам. То же касается и геномов клеток. Здесь
прослеживаются филы родственных организмов. Цифрами обозначены одногенные
вирусы. Буквами – многогенные вирусы и клетки растений. Клетка g является
родственником всем остальным клеткам и вирусам, потому что содержит их гены.
Механизм, регулирующий добавление новых генов в молекулу ДНК известен –
рекомбинация (коньюгация) молекул ДНК путем вирусного заражения. Методом
выращивания наноколоний молекул РНК на геле [6, 41, 42] было
экспериментально доказано, что в популяциях молекул РНК свободно происходит их
рекомбинация (коньюгация) с образованием химерных молекул РНК, образовавшихся
путем соединения двух или более молекул РНК. Молекулы РНК также могли
обмениваться своими участками. Так могли возникнуть самые разные комбинации
генотипов РНК уже в начале их эволюции.
Естественно, тем же путем могла происходить и коньюгация молекул ДНК у
первичных вирусов, результатом которой является избыточный набор генов.
Коньюгация вирусов должна существовать, потому что есть малогенные вирусы и есть
многогенные. Если в первичных малогенных вирусах (прото-клетках) молекула РНК/ДНК
появилась как результат неких химических реакций на матрицах тех или иных
кристаллов (глин и т.д., здесь не будем рассматривать все подробности теории
мира РНК), то откуда появились добавочные и весьма многочисленные и
разнообразные гены в остальных генерациях многогенных вирусов? Ответ:
многочисленные и разнообразные гены в остальных генерациях многогенных вирусов
появились только в результате срабатывания механизма генной мультипликации.
Видов вирусов чрезвычайно много, это наиболее многочисленные организмы на
Земле (до 250 миллионов в одном миллилитре воды). Они различаются между собой
свойствами, которые зависят не только от строения их генов (вида генов), но и от
числа генов в каждом вирусе – от 1-го до нескольких десятков, сотен и даже тысяч
генов. Многогенные вирусы могли появиться только одним путем – путем коньюгации.
Из одногенных вирусов путем коньюгации их молекул ДНК получились многогенные
вирусы, из которых в дальнейшем произошли первые клетки – прокариоты.
Присоединение новых генов к геному создает генную избыточность. Если жертва
выживает, это значит, что она не только сумела неким образом подавить
деятельность генома вируса, иначе она бы не выжила, но и приобрела новые гены,
внедренные в её геном вируса.
У каждой клетки есть собственные биологические конструкции, которые она может
синтезировать, и для этого у неё есть необходимые для этого действующие генные
наборы. После такого заражения, кроме собственных необходимых ей действующих
генов, у неё появляется и лишний недействующий («мусорный») заблокированный
вирусный ген или гены. Если их не заблокировать, то они будут размножать
вирусные копии и клетка погибнет (лизис). Но если их заблокировать, то
появляется генная избыточность (лизогения). Так работает механизм генной
мультипликации, основанный на вирусном заражении.
Этот механизм наиболее эффективно работает у простейших организмов – вирусов,
прокариот, низших растений и т.д., потому что у них не развита иммунная система,
противодействующая вирусному заражению. Мир вирусов и бактерий продолжался на
Земле более 3-х с половиной миллиардов лет и за это время механизм генной
мультипликации путем вирусного заражения накопил огромное число генов в их
геномах. Это и проявилось в эдиакарском и кембрийском взрывах более
полумиллиарда лет назад, когда как бы вдруг появилось множество самых
разнообразных многоклеточных организмов. Но это было не вдруг, потому что
одноклеточные за 3 миллиарда лет накопили множество генов (избыточность генов).
А когда в атмосфере Земли появился свободный кислород, то появилось много
аэробной энергии и сразу же (это «сразу» продолжалось несколько миллионов лет)
появилось множество многоклеточных организмов, которые раньше не могли быть
из-за недостатка энергии для своих действий. Тригерром запуска «спящих» генов
явилось повышенное содержание кислорода в атмосфере и гидросфере Земли.
Благодаря вирусному заражению (генной мультипликации) появилось очень большое
количество видов живых организмов, имеющих очень «богатые» геномы с большой
генной избыточностью. По мере последующих вирусных заражений и накопления генов
в геноме появлялись полноценные (комплектные) наборы генов, способные управлять
синтезом многих биологических конструкций. Эти наборы генов «спят» до поры до
времени и если возникает необходимость в этих конструкциях, то эти наборы генов
«пробуждаются» и начинают действовать, синтезируя новые биологические
конструкции. И тогда как бы вдруг появляется новый вид. Таким образом появление
новых видов во многом обязано вирусному заражению.
Механизм генной мультипликации, это механизм появления нового вида за время
жизни одной особи – материнской особи, у которой уже был большой комплект
нажитых её предками «мусорных» генов от предшествующих вирусных заражений. В
некий момент некий её предок заразился очередным вирусом и получил «последний»
недостающий ген для синтеза некого нового органа (эффект «соломинки с хребтом
верблюда»). Чтобы выжить, этот предок блокировал этот ген и продолжал жить,
выполняя свою основную цель – размножался. Сам этот предок ещё принадлежал
прежнему виду, но его непосредственный потомок и все остальные внутчатые потомки
уже могли принадлежать новому виду (могли, но ещё не принадлежали!), потому что
они наследовали уже новый полноценный набор генов, которые могут начать
действовать и управлять синтезом элементов новых органов. Но до поры до времени
эти гены у них были в «спящем состоянии» и находились в недействующей части
генома («мусорные гены»). Все биохимические циклы, управляемые действующей
частью генома у них те же, что и в материнском виде, иначе различие
биохимических циклов привело бы к анатомическому различию особей в каждом
биологическом виде. Поэтому предки данной особи, включая её самую, могли
скрещиваться с материнским видом и размножаться. Но эти «спящие» гены могут
«проснуться». И тогда у особей-потомков могут начать действовать новые
биохимические циклы, которых нет у матери. В этом случае скрещивание
особи-потомка с особями материнского рода невозможно, потому что они будут
принадлежать новому виду. Но пока они не «проснулись», материнский вид
продолжает размножаться, увеличивая число потомков с новым геномом. И в некий
определённый момент срабатывает некий триггер и рождается множество особей
нового биологического вида.
Так за время жизни одной особи появляется новый вид – несколько
особей-матерей рожают особи нового вида, которые «отпочковываются» от
материнского рода и уже с ним не пересекаются (новая фила не скрещивается со
старой филой). А поскольку множество таких матерей рожают множество особей
нового вида, то для них проблемы репродуктивной изоляции не существует.
Эволюционный прирост нормальной, а не мутированной молекулы ДНК – это
нормальный процесс её изменения за счет прироста новых генов путем вирусного
заражения, приводящий к её удлинению и созданию достаточной избыточности генов
для появления полноценных НАБОРОВ генов, что может дать положительный
эволюционный эффект. Так работает рекомбинантный механизм вирусного заражения
(механизм генной мультипликации) для наращивания генной избыточности в молекуле
ДНК.
Перенос генов даже между не родственными организмами – это известный факт.
Ему предшествовал механизм вирусного заражения. Этот механизм переноса имеет
множество рекомбинантных компонентов и широко используется вирусами для своего
размножения в клетках других организмов. Вирусы (фаги) «встраивают» свой геном в
ДНК клетки хозяина и так они размножаются. Этот же механизм является основным
для наращивания генов в геномах всех живых организмов (механизм генной
мультипликации путем вирусного заражения).
Да, в результате вирусного заражения носитель вируса (зараженный организм)
может и погибнуть. Но если он не погибает, то сохраняет новые встроенные
дополнительные вирусные гены и передает их по наследству своим потомкам. Выжить
и не погибнуть зараженный организм хозяина может лишь в том случае, если он
сможет заблокировать новые и, в данном случае, балластные («мусорные») вирусные
гены, чтобы их функция до поры до времени не конкурировала с другими генами
клетки-хозяина. Когда же таких балластных («мусорных») генов наберется
достаточно, чтобы их совместная функция могла привести к синтезу новой
конструкции (полный комплект генов), дающей положительный эволюционный эффект,
они из балластных могут превратиться в действующие. И тогда, как бы вдруг,
появится новый вид, хотя процесс набора полного комплекта новых генов занимает
многие миллионы, а то и миллиарды лет периодических вирусных заражений.
Механизм блокирования активности «мусорных генов» так же известен – это
механизм РНК-интерференции, процесс подавления экспрессии гена (передачи
наследственной информации от гена на РНК или белок) на стадии транскрипции,
трансляции, деаденилирования или деградации мРНК при помощи малых молекул РНК.
РНК-интерференция обнаружена в клетках многих эукариот, в том числе у животных,
растений и грибов. Система РНК-интерференции играет важную роль в защите клеток
от паразитирующих генов — транспозонов и вирусов, а также в регуляции развития,
дифференцировки и экспрессии генов организма. Возможны и другие механизмы (piRNA,
различные антисмысловые РНК и прочее), которые рассматривать здесь не будем.
Важен сам факт – нам известен механизм блокады «мусорных» генов.
Несмотря на все различие между минеральными (неживыми) и биотическими
(живыми) системами их объединяет одно общее – они все управляются единой
иерархией законов природы, которая включает в себя законы мира минералов и
законы биотических миров, в том числе и законы органической и биохимиии,
биологии и т.д.[36] Поэтому нет ничего необычного в том, что
первоначально коды жизни были «записаны» в условиях синтеза молекул РНК и ДНК,
когда они синтезировались на матрицах кристаллов (или неких глин), если верна
гипотеза о мире РНК. Другими словами, условия синтеза на матрицах глин были
переведены в записи на молекулах РНК и ДНК. Несмотря на очень большое число
кодов, это число настолько же конечно, насколько конечно число условий синтеза
молекул РНК и ДНК. Сами же молекулы РНК и ДНК так же являются минеральными
образованиями, т.е., «особями» мира минералов (они не живые).
Следовательно, минеральный мир обусловил появление систем мира РНК и, в
последующем, систем всех остальных биотических миров. Появление Солнечной
системы, планеты Земля, океанов на ней и вся минеральная и биологическая история
Земли – это все звенья одной цепи последовательных действий иерархии законов
природы, целью которой было появление и развитие жизни на Земле и, возможно, на
других планетах (антропогенез).[36]
Записи кодов различных генов «разбросаны» по всей Земле в виде кодов геномов
вирусов, которых великое множество. То, что в каждом мл морской воды находится
до 250 миллионов вирусов и не все они фаги указывает на то, что «минеральное»
рождение вирусов (прото-клеток) все ещё продолжается и сегодня. Если бы все они
были бы фагами (размножались бы в других живых организмах), то морская вода была
бы стерильна от всех живых организмов. Да и все вирусы должны были бы исчезнуть
ввиду того, что они же «съели» бы своих жертв. Такое огромное число вирусов в
морской воде говорит об их «минеральном» происхождении.
Если их столько содержится в одном кубике морской воды, то сколько же их
«разбросано» по всей Земле? Если переводом триплетных кодов в геномы вирусов
занимались системы минерального мира (кристаллы аппатита, условия синтеза
молекул РНК и ДНК и т.д.), то переводом этих кодов в записи уже на молекулы РНК
и ДНК всех остальных биосистем занимались сами вирусы. Вирусы стали объединять
свои геномы и получались сначала сложные составные прото-клетки (многогенные
вирусы). А затем, когда набирался полный комплект генов, способных управлять
синтезом клеточных мембран и других клеточных органелл, появлялись полноценные
клетки. А в этих клетках, как и в клетках всех остальных живых систем, прирост
числа генов продолжался за счет того же рекомбинантного механизма вирусного
заражения для наращивания генной избыточности в их молекулах ДНК, если
клетки-хозяева выживали после такого заражения. Так получались клетки с
избыточным набором генов.
Следует отметить и ещё один аспект генной мультипликации, основанной на
вирусном заражении – вирусы-фаги всегда специфичны только для определённых живых
организмов, причём очень и очень избирательны. Это говорит о том, что геном
вируса-фага подходит для другого генома только (!) определённого вида. Но это же
значит, что вирусы-фаги как бы подыскивают только определённые геномы и
встраивают в них свои гены. А это и есть целенаправленная генная мультипликация!
Создается такое впечатление, что гены, как кубики, рассыпаны по всей Земле, но
эти «кубики» сами знают, к каким другим «кубикам» они подходят и ищут их для
воссоединения с ними путем вирусного заражения. Отсюда, генная мультипликация –
это не случайный, а целенаправленный процесс!
Вирусное заражение является основным механизмом обогащения генома на всех
уровнях развития биологических систем, начиная с одногенных вирусов и заканчивая
организмом человека, создавая его генную избыточность. Генная избыточность
появилась только за счет последовательного захвата очередных ДНК прото-клеток
(последовательного захвата очередных генов вирусов – вирусная конъюгация),
происходящего при последовательном вирусном заражении.
Это насильственное взаимное обогащение геномов путем вирусного заражения
(конъюгации) происходит под управлением иерархии законов природы (законов химии,
биохимии и генетики). И даже в последующем, когда вирусы заражали все более
сложные многогенные биосистемы и присоединяли свои гены к геномам заражаемых
многоклеточных, это тоже было насильственным обогащением этих геномов,
навязанным биологической (в частности биохимической) ветвью иерархии законов
природы.
Такое насильственное обогащение геномов за счет вирусного заражения всех
биосистем продолжается и сегодня, потому что существуют вирусные заражения и
болезни, связанные с этим. И так продолжается вплоть до уровня Homo sapiens’а,
но не выше, потому что на Homo sapiens заканчивается биологическая эволюция и
начинается надбиологическая (социально-технологическая) эволюция. Далее если и
будет продолжение генной мультипликации, это будет управляемая человеком генная
мультипликация (технологическая-медицинская, а не биологическая).
Следы различных вирусных геномов были обнаружены у многих млекопитающихся.
Было показано, что за последние 40 миллионов лет, почти половина из 48 изученных
видов позвоночных приобрела последовательности, связанные с генами двух вирусных
семей – Ebola/Marburgviruses и Bornaviruses, которые вызывают смертельные
геморрагические лихорадки и неврологическую болезнь, соответственно.[28]
В данном случае позвоночные заплатили очень дорогую цену за приобретение
дополнительных генов. Но, видимо, цена достойна приобретению: да, многие
организмы погибли, но в итоге появились новые более развитые и жизнестойкие
биологические виды с новыми наборами генов (рис. 4).
![Рис. 4. Филогенетическое дерево позвоночных животных, в которых были обнаружены геномы Ebola/Marburgviruses, и Bornaviruses. Mya – миллионов лет назад.[28]](images/art/all20/new_pa5.gif)
Рис. 4. Филогенетическое дерево позвоночных животных, в которых были
обнаружены геномы Ebola/Marburgviruses, и Bornaviruses.
Mya – миллионов лет назад.[28]
Если бы вирусные гены были совершенно индефферентны в биологическом аспекте,
то такие гены были бы понастоящему мусорными. То, что многие вирусные
заболевания чрезвычайно опасны, говорит о том, что белки, синтезируемые этими
новыми внедренными генами, обладают очень высокой биологической активностью. Эти
белки могут активно вмешиваться в биохимические циклы клеток-хозяев, отравляя и
даже убивая их. Но клетки-хозяева не всегда гибнут, потому что блокируют вновь
внедрённые гены, превращая их в мусорные. И таких мусорных генов может набраться
достаточно, чтобы синтезировать элементы новых биологических конструкций. При
«умелом» использовании вновь приобретенных генов можно получать очень высокий
биологический эффект. Острым и опасным инструментом можно легко пораниться, но
только с ним можно сделать некую специфическую работу. Поскольку каждый из них в
отдельности биологически активен, то при удачном сочетании такой набор генов
может контролировать синтез очень полезных биологических конструкций. Именно
этот принцип и лежит в основе генной мультапликации путем вирусного заражения.
40 миллионов лет составляют около 1% от времени эволюции жизни на Земле. Если
бы прирост числа генов всегда происходил с такой скоростью, то самая сложная
молекула ДНК включала бы не более нескольких сот генов и времени эволюции не
хватило бы на постройку геномов не только человека, но и более простых клеточных
форм жизни вообще. Однако, чем дальше назад по времени, тем биосистемы были
проще, и, следовательно, эффективность средств антивирусной защиты у живых
систем была ниже, тем легче вирусы «вписывали» свой геном в ДНК клетки хозяина и
тем интенсивнее был процесс вирусного наращивания генов (генной мультипликации)
в молекулах ДНК клеток-хозяев. К этому следует добавить, что процесс накопления
генов одноклеточными (простейшими бионтами) продолжался более 3-х миллиардов
лет. Можно представить себе, сколько генной избыточности накопилось за это
время.
В результате накопления «мусорных» генов может получиться, а может и не
получиться полноценный набор генов, способный синтезировать какую-либо полезную
для данной особи биологическую конструкцию. Если полноценный набор генов
получается и если в такой конструкции появится необходимость, то по закону
Дарвина клетка может ответить неблагоприятным условиям внешней среды синтезом
соответствующих белков, из которых будет построена необходимая биологическая
конструкция. Так путем вирусного заражения формируется генная избыточность
биологических особей, приводящая, в конечном итоге, к появлению новых
биологических конструкций, что определяет новые свойства особей и появление
нового вида. Закон Дарвина при этом выполняет не видообразующую роль, а
фильтрующую (селективную) роль, выбирая из всего множества вновь образующихся
видов только те, которые наиболее оптимально подходят для условий именно данной
внешней среды.
Слияния же самих межвидовых геномов одно- и многоклеточных не происходит.
Этому препятствует различие биохимических циклов, управляемых разными
действующими частями геномов (у разных видов разные действующие части геномов).
Поэтому филы не перекрещиваются. Так за счет вирусного заражения и накопления
генной избыточности происходит эволюция биосистем.
Такое взаимодействие – сотрудничество нескольких (в минимуме – двух)
прото-клеток (вирусов) являлось первым примитивным симбиозом. Симбиозом потому,
что разрушения их молекул ДНК не происходило, в то время как их общий геном
усиливался, приобретая дополнительные свойства. Благодаря этому была заложена
основа для появления уже настоящих клеток, содержащих сначала тысячи, а затем и
многие десятки тысяч генов в симбиотически рекомбинированных молекулах ДНК для
производства всего комплекса белков внутри- и внеклеточных конструкций. Этот же
вирусный рекомбинантный механизм наращивания генов сохранился на всех
последующих уровнях развития живых систем.
Первые прото-клетки (вирусы) принадлежали миру РНК. Они были продуцентами,
потому что продуцировали гены и белок, а в качестве метаболитов использовали
системы (атомы и молекулы) лежащего ниже их минерального мира (нуклеотиды и
аминокислоты). Все виды системных взаимодействий более сложных биосистем
(сапрофитов, редуцентов, паразитов и хищников) подразумевают наличие системы
пищеварения, которой у вирусов нет, потому что у них нет цитоплазмы, содержащей
биохимические котлы, и цито-плазматической мембраны, отделяющей цитоплазму от
внешней среды. Поэтому среди них не может быть и истинных сапрофитов, паразитов
и хищников. Но вирусы могут использовать систему пищеварения клеток-хозяев.
Сегодняшние вирусы – это опосредствованные (непрямые) сапрофиты, паразиты и
хищники, в которых они превратились за 4-4.5 миллиарда лет существования и
взаимодействий с другими прото-клетками и живыми системами более высоких уровней
сложности.
Между собой вирусы взаимодействуют как хищники-симбионты, объединяя свои гены
в один общий геном. Хищники потому, что стремятся захватить геном другого
вируса, а симбионты потому, что не разрушают захваченные геномы (гены), а
объединяют их. Вирусные симбионты определили появление многогенных вирусов, а
через них и появление уже всех остальных живых систем более высоких уровней
сложности. Слияние двух или более молекул вирусных ДНК увеличивает их
генетическую емкость и создает генную избыточность, которая, в конечном итоге,
приводит к появлению новых биологических конструкций в особях и появлению новых
биологических видов.
У высокоразвитых организмов выздоровление означает не только появление
антивирусных иммунных антител в крови, но и способность выжившего организма
блокировать вирусные гены, которые все таки были встроены в их геном в
результате данного заражения. В таких случаях вирус уже не может эффективно
размножаться, используя геном этих особей, потому что их организм умеет
блокировать гены этих вирусов. И это может быть настолько эффективной защитой,
что даже не требуется антител иммунной системы особи для подавления размножения
вирусов.
В таких случаях заражённая особь превращается в вирусоносителя. В её
организме нет иммунных антител или их титр низок. В таких условияъ вирус может
размножаться, используя для этого её метаболиты, размножаясь в небольших
количествах и не составляя ей метаболическую конкуренцию (лизогения,
псевдолизогения и хроническая инфекция). Но он не может использовать уже
внедренные в геном клетки-хозяина его (вирусные) гены, потому что они
блокированы. Эффективность такого размножения этих вирусов (без использования
генома клеток хозяина) настолько низкая, что вирус не представляет опасности для
данной особи, её организм не борется с ним и вирус спокойно проживает в ней,
паразитируя. По этой же причине особи, уже перенесшие такое вирусное заражение,
могут оказаться вообще не восприимчивыми к повторным заражениям данным вирусом.
Поэтому и их потомки также могут оказаться невосприимчивыми к таким вирусным
заражениям, даже без наличия соответствующих антител в крови, потому что
унаследуют от своих родителей как новые вирусные гены, так и способы их
блокировки. Многочисленные случаи вирусоносительства говорят об этом.
Таким образом, любое вирусное заражение любых живых организмов приводит не
только к заболеванию, которое может закончиться смертью (лизис) или
выздоровлением, но и к увеличению генной избыточности, если заражение
заканчивалось выживанием (лизогенией). Если наступала смерть, геном данной особи
выбывал из эволюции. Но если наступало выздоровление, то геном данной особи
обогащался ещё одним геном, увеличивающим его генную избыточность и дающим ещё
один шанс эволюционной устойчивости. Поэтому в прокариотах продолжился процесс
накопления генной избыточности и это привело ко всему разнообразию современных
видов живых систем.
И это именно избыточность, так как мы знаем, что только небольшая часть генов
любого генома участвует в синтезе тех или иных белков. Например, у человека
только 1.5% генома состоит из кодирующих белок экзонов.[31]
Следовательно, геном любой биологической системы содержит действующие
(кодирующие белок экзоны) и недействующие (избыточные) гены. Причем, для синтеза
любой биологической конструкции необходим не один ген, а целый набор генов,
потому что для постройки, например, носа требуются кости, хрящи и несколько
типов мягких тканей. Даже для постройки простейшего двигателя (протоно-двигателя)
жгутика бактерии требуются 40 белков, для синтеза которых требуются 40 генов и
плюс ещё пару десятков генов для синтеза остальных механизмов, которые собирают
этот двигатель и встраивают его в мембрану бактерий. Естественно, биологические
конструкции более развитых живых систем требуют более многочисленных наборов
генов.
Доля мусорных генов в геноме особи различна и зависит от уровня развития
данной особи. Бактерии E.coli накопили около 4200 генов и из них только 62%
кодируют используемые бактерией белки.[14] У человека в геноме
несколько десятков тысяч генов и плюс ещё несколько сот тысяч генов, если
учитывать механизм сплайсинга, но из них используется всего 1.5%.
Вывод: чем больше геном, тем больше «мусорных» генов.
Со временем, особенно после появления многоклеточных и развития их иммунных
систем, вирусное заражение все затруднялось. Поэтому у развитых организмов
заражение происходит только от очень вирулентных (сильно заразных), способных
преодолевать сопротивление иммунной системы и поэтому очень опасных вирусов. Но
даже если заражение и происходит, это не всегда означает передачу новых вирусных
генов своему потомству по двум причинам:
- у многоклеточных стала развиваться система иммунной защиты,
противодействующая вирусному заражению, тем более эффективная, чем более
сложным был многоклеточный организм,
- чтобы у многоклеточных могла осуществиться передача новых вирусных генов
потомству, должно быть заражение не любых, а именно половых клеток
(сперматозоидов и/или яйцеклеток).
Поэтому, по мере развития биосистем скорость наращивания генома вирусными
генами за счёт вирусного заражения замедлялась по экспоненте и у высших
многоклеточных биосистем, включая человека, она чрезвычайно медленная.
Число видов вирусов огромно и среди них всегда можно найти те или иные ДНК,
способные производить те белки, которые могут встречаться у тех или иных
биосистем, независимо от их уровня сложности. Другими словами, коды любых белков
записаны в геномах всего разнообразия вирусов. За 3-4 миллиарда лет эволюции
многие из этих кодов тем или иным путем сгруппировались в виде различных молекул
ДНК, составив геномы различных живых организмов. Все это говорит лишь о том, что
в течение очень длительных промежутков времени в молекулах РНК, а в дальнейшем и
ДНК, путем генной мультипликации создавалась генная избыточность.
Низко развитые организмы потому и остались низко развитыми, потому что ещё не
успели накопить генную избыточность вирусного происхождения, подобную высоко
развитым организмам. Иначе как объяснить сегодняшнее существование примитивных
организмов, включая вирусы и бактерии. Не накопил генов, не развился и остался
на прежнем уровне. Этот вывод так же указывает на то, что добавление нового гена
в геном – это достаточно редкое событие.
Очевидно, прирост числа вирусных генов в молекуле ДНК за счёт вирусного
заражения происходит по экспоненте и сегодня этот процесс уже значительно
замедлен. Но он ещё продолжается, потому что есть ещё множество вирусных
заболеваний. С учетом прежних больших скоростей такого прироста генов в молекуле
ДНК, времени эволюции должно было хватить для появления генома человека. Все это
дает основание утверждать, что геномы всех клеток растений, животных и человека
состоят из генов вирусов, набранных за время эволюции.
Но в силу замедления прироста вирусных генов за счёт вирусного заражения,
появился новый механизм наращивания вирусных генов в геномах сложных организмов
– механизм полового скрещивания (половое размножение – секс).
И если при вирусном заражении в основе генной избыточности работают два
механизма:
- объединение генов различных вирусов в один общий геном, приводящее к
появлению сложных многогенных вирусов и прокариотов (коньюгация вирусных
ДНК),
- повторные вирусные заражения прокариотов и всех остальных живых систем
любой степени сложности с продолжением встраивания вирусных генов в их геном
(генная мультипликация),
- которые по мере развития иммунных систем заражаемых организмов стало
осуществлять все труднее и труднее, то появление ещё одного механизма
(процесса), который также позволяет обогащать генную избыточность, значительно
облегчило эту задачу.
На Земле проживает весьма многочисленное множество особей (сотни миллиардов)
самых различных биологических видов, содержащих геномы с очень большим
разнообразием «мусорных» генов, состоящих из очень большого разнообразия
остатков вирусных генов. Если неким образом совместить «мусорные» части геномов
различных особей и сделать механизм, дополняющий совмещённые части каждой из ДНК
«мусорными» генами из геномов половых партнёров, то можно получить обогащённые
«мусорные» части геномов.
Этим механизмом является механизм полового скрещивания. В литературе принято
несколько иное название этого явления – половое размножение.
Первые организмы, возможно размножавшиеся половым путем, относятся к
стенийскому периоду мезопротерозоя (около 1-го миллиарда лет назад). Образование
половых клеток, как правило, связано с прохождением мейоза на какой-либо стадии
жизненного цикла организма. В большинстве случаев половое размножение
сопровождается слиянием половых клеток (гамет). При этом восстанавливается
удвоенный относительно гамет набор хромосом. В зависимости от систематического
положения эукариотических организмов половое размножение имеет свои особенности,
но, как правило, оно позволяет объединять генетический материал от двух
родительских организмов и позволяет получить потомков с комбинацией свойств,
отсутствующих у родительских форм.
Эффективности комбинирования генетического материала у потомков, полученных в
результате полового размножения, способствуют:
- случайная встреча двух гамет;
- случайное расположение и расхождение к полюсам деления гомологичных
хромосом при мейозе;
- кроссинговер между гомологичными хромосомами.
Термин «половое скрещивание» более верно отображает суть явления, чем термин
«половое размножение». Скрещиваются не полноценные молекулы ДНК, а их продольные
половинки, от каждого родителя по одной. Кроме того, термин «половое
размножение» вносит некую неопределённость. Партеногенез не предусматривает
слияния гамет (продольных половинок молекул ДНК от отца и матери), хотя и
предполагает деление неоплодотворенных яйцеклеток (размножение половых клеток).
Сперматозоиды же никогда не делятся
Хотя партеногенетическое размножение не предусматривает слияния мужских и
женских гамет, партеногенез всё же считается половым размножением только по той
причине, что новый организм развивается из половой клетки (ооцита). Считается,
что партеногенез возник в процессе эволюции раздельнополых форм.
В тех случаях, когда партеногенетические виды представлены (всегда или
периодически) только самками, одно из главных биологических преимуществ
партеногенеза заключается в ускорении темпа размножения вида, так как все особи
подобных видов способны оставить потомство. Такой способ размножения встречается
у некоторых животных, хотя чаще среди относительно примитивных организмов.
Партеногенез относят к половому размножению и отличают от бесполого
размножения, которое осуществляется при помощи соматических органов и клеток
(размножение делением, почкованием и т. п.). Но, по большому счету,
оплодотворенная половая клетка является обычной клеткой, похожей на все
остальные. Отличие от остальных клеток только в том, что у неё разблокированы те
гены, которые заблокированы у дифференцированных клеток, и заблокированы все
остальные гены, которые разблокированы в той или иной степени у
дифференцированных клеток. Поэтому ооциты, вообще говоря, являются главными
стволовыми клетками, у которых по мере их дифференциации блокируются или
разблокируются те или иные гены. Поэтому партеногенез половым размножением не
является. Это несколько необычный, но по механизму рядовой способ бесполого
размножения типа почкования.
Половое скрещивание принципиально отличается от партеногенеза. При
скрещивании происходит дополнение генома одного родителя генами другого родителя
(к одной половинке молекулы ДНК одного родителя присоединяется другая половинка
другого родителя). При партеногенезе никакого дополнения генами нет. Поэтому
термин «половое скрещивание» полностью отображает сущность происходящего
процесса – геномы родителей скрещиваются.
Для чего вообще существует половое скрещивание? Какой смысл в скрещивании
двух молекул ДНК от мужской и женской особей, если их действующие наборы генов
совершенно одинаковы? И почему скрещиваются именно половинки вдоль расщепленных
молекул ДНК (по одной из спиралей от каждого родителя из двойной спирали ДНК)?
Ведь это, на первый взгляд, лишняя и очень сложная работа – расплести из
гексонов молекулу ДНК, расщепить её на две половинки, затем синтезировать только
одну половинку и только после этого упрятать эту половинку (одинарную спираль
молекулы ДНК) в сперматозоид или в яйцеклетку. И всё это поставить в очень
проблематичную зависимость от случайности встречи половых партнёров. И почему-то
этот процесс наблюдается практически у всех многоклеточных и в общих чертах
одинаков.
На вероятный ответ указывают два факта:
- молекула ДНК состоит из двух совмещенных спиралей, а не из одной спирали
ДНК,
- при половом скрещивании соединяются отдельные продольные половинки
молекулы ДНК, по одной от каждого родителя.
Если действующие наборы генов родителей одинаковы, например, содержат
одинаково нормальные гены, то при скрещивании с генами ничего не происходит и
плод будет нормальным. Если они содержат одинаковые дефекты генов, то при
скрещивании с ними также ничего не происходит, но плод родится таким же
дефектным, как и его родители. Если они имеют разные дефекты, то при
оплодотворении будет исправление генных дефектов и плод родится нормальным (рис.
5).
При соединении двух одинарных спиралей в одну двойную, там, где есть разница
в содержании генов в материнской и отцовской одинарных спиралях (молекула ДНК
окажется дефектной), на месте несимметричного дефекта будут своеобразные дыры.
Если у одного из родителей в этих местах генные наборы нормальны, то происходит
исправление дефектных по образу и подобию генов здорового родителя.
Этими «дырами» могут быть, например, деструктивные мутации в действующей
части каждой половинки генома, причем в каждой из них разные. При оплодотворении
репликационный аппарат яйцеклетки достроит недостающие части, заполняя дефекты,
например, материнской половинки ДНК тем, что содержится на отцовской половинке
ДНК, исправляя «ошибки». И наоборот. В итоге этот аппарат заполнит дефекты
двойной спирали молекулы ДНК оплодотворенной яйцеклетки и в итоге у плода будет
полноценная молекула ДНК, содержащая нормальные гены матери и гены отца.
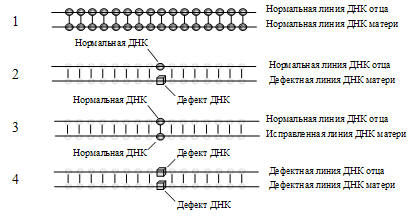
Рис. 5. Образование двойной спирали молекулы ДНК плода
из линий половинок молекул ДНК отца и матери.
1 – слияние нормальных линий отца и матери; 2 – слияние нормальной линии
ДНК отца с дефектной линией ДНК матери (образуется генный дефект ДНК); 3 –
исправление дефекта линии ДНК матери через образ и подобие соответствующего
участка линии ДНК отца (генный дефект исчез); 4 – закрепление генного
дефекта ДНК у близкородственных родителей с одинаковыми дефектами ДНК.
Есть определённый биологический смысл полового скрещивания и «мусорных»
генов, которые у мужской и женской особей даже одного биологического вида
разные. Это значит, что при половом скрещивании геном плода получит «мусорные»
гены и матери, и отца. В результате такого скрещивания геном будет содержать
больше генов («мусорных» генов), чем каждый геном его матери и отца по
отдельности, потому что он будет включать в себя те «мусорные» гены, которые
есть у матери и нет у отца, и те «мусорные» гены, которые есть у отца и нет у
матери, включая те гены, которые есть и у отца, и у матери.
Таким образом, за счет полового скрещивания происходит процесс исправления
генетических нарушений и процесс накопления «мусорных» генов. При этом генная
избыточность нарастает за счёт новых вирусных генов, но без излишнего вирусного
заражения, чреватого ненужной смертностью. Потомок приобретает и новые вирусные
гены от родителей, не заболевая при этом, и исправленые гены, нарушенные генной
деструкцией.
Поэтому есть определённый биологический смысл в скрещивании, которое
исправляет генетические дефекты и накапливает генную избыточность. Тот же
механизм, который достраивает дыры в «мусорной» части молекулы ДНК, может
исправлять дефекты в действующей части генома, сравнивая обе половинки в
оплодотверенной яйцеклетке. В двойной спирали ДНК есть возможность исправлять
правильность кода. Поэтому у отца и у матери все молекулы ДНК построены в виде
двойной спирали, а в половых клетках содержатся только одинарные спирали.
Потому и существует природный запрет на близкородственные браки
(кровосмесительство и инцест), где геномы больше похожи на клоны. Чем дальше
родство особей, тем меньше шансов на одинаковые дефекты (мутации) генов, тем
больше шансов на исправление генов того родителя, у которого они дефектные. При
этом у потомка появляется больше шансов на различие состава «мусорных» генов.
Чем больше это различие, тем больше шансов на приобретение потомком новых генов,
тем больше шансов на появление нового более совершенного вида.
Вероятно поэтому природой сделаны такие пышные хвосты павлинов, развесистые
рога оленей, такие огромные шипы и щитки у динозавров, такие висячие и
переливающиеся многоцветьем сосульки на головах индюков и прочие совершенно
ненужные «в хозяйстве» украшения, чтобы показать самке все богатство своих
генов. Чем богаче геном, тем больше всяческих «ненужностей» в украшениях райских
птичек и прочей живности, которые не играют никакой биологической роли, кроме
как для использования в брачных играх и демонстрации своего генного богатства.
Если у меня есть множество всяческих «ненужностей», то у меня есть очень богатый
геном и, следовательно, очень богатое генное наследство, которое, как мы знаем,
заключено в его «мусорной» части.
Для самого спаривания и зачатия сами половые игры не нужны. Чего там,
пробегал мимо, спарился и побежал дальше. У всех растений и у большинства низко
развитых животных так и происходит. Ни в какие игры они не играют, а пользуются
случаем. Это происходит, вероятно, потому, что у них иммунная система слабая.
Поэтому вирусное заражение у них протекает легче (с точки зрения самого
заражения, но не с точки зрения выживаемости при заражении) и для обогащения
генома оно имеет бóльшее значение, чем половое скрещивание.
В половые игры играют, как правило, уже достаточно развитые животные. Зачем
нужны половые игры, которые кроме осложнений в жизни на первый взгляд ничего не
дают? Слишком много ненужных хлопот и, более того, слишком много риска для жизни
играющих. Тем не менее сам факт существования половых игр, как правило, очень
многоэтапных и сложных, говорит о некой их биологической целесообразности. И эта
целесообразность заключается в демонстрации собственного богатства генов,
включая «мусорные», и хорошей настройки действующих генов.
Для нас всегда было загадкой, для чего лосям и оленям такие рога, ведь
таскать такую тяжесть на голове тяжело и неудобно. Если уж для защиты, то не
лучше ли маленькие и острые, как у лани? Но тогда нечего будет показывать своим
самкам! Есть такие, которые вообще не дерутся, а воняют и у них нет никаких
излишних украшений. Чем больше воняет, тем он для самки предпочтительнее. Многие
птички устраивают только демонстрационный конкурс – чем лучше расцветка
оперения, тем больше генное богатство «мусорной» части ДНК. В брачных же
бойцовских турнирах демонстрируется качество настройки генов действующей части
генома. У кого настройка лучше, тот сильнее, ловчее, удачливее, красивее (с
точки зрения брачующихся), вонючее и голос его громче или заливистее и т.д. А
самка хоть и выбирает наиболее выдающегося ухажера, но это не значит, что она
четко осознает то, что она делает, потому что у неё нет никакого понятия о ДНК и
генах. Все за неё делает её инстинкт, который откуда-то все это знает. Откуда?
Эти знания в него (в инстинкт) заложены иерархией законов природы.
Бóльшая дальность родства способствует исправлению генных ошибок в
действующей части генома потому, что нарушения генов в геноме одной персоны не
могут быть такими же, что в геноме другой персоны из-за низкой вероятности
одинаковых нарушений у двух далеко родственных особей одного вида. Чем дальше
родство, тем меньше шансов на одинаковость генных нарушений. При половом
сведении двух половинок спирали молекул ДНК механизм репликации может исправить
нарушенные участки одной половинки спирали ДНК по образу и подобию ненарушенного
участка второй половинки молекулы ДНК. Возможно потому инцест дает большой
процент генных нарушений, а дальнородственные браки сохранили наш геном на
протяжении многих миллионов лет.
Такой механизм исправления генетических нарушений объясняет явление
импритинга, наблюдающийся в природе. Термин «импринтинг» впервые предложил
американский генетик X. Кроуз (Колумбийский университет) в 1960 году для
описания элиминации отцовских хромосом у насекомых. Геномный импринтинг
подчеркивает факт, что наследуются изменения генной активности, обусловленные
различным происхождением хромосом (от отца или матери), а не структурными
перестройками генетического материала. Было установлено, что в отдельных
участках хромосом, которые подвержены геномному импринтингу, экспрессируется
только один отцовский или один материнский аллель в отличие от обычной
диаллельной экспрессии генов. Если геномному импринтингу подвержен материнский
ген, то экспрессируется только отцовский аллель и наоборот.[ 3, 10, 18, 25,
30 и другие]
Существует особый класс болезней – «болезни геномного импринтинга». Их
количество превышает 40 нозологических форм (синдромы Прадера-Вилли, Апгельмана
и другие). Природа синдрома Прадера-Вилли связана: в 75% случаев с делецией
критического района хромосомы 15 отцовского происхождения, в 20% –
однородительской дисомией и примерно в 5% – мутациями «импринтинга»; если же
делеции подвергается тот же участок хромосомы 15 материнского происхождения, то
в 75% случаев появляется заболевание с другой, отличной от синдрома
Прадера-Вилли, клинической картиной – развивается синдром Ангельмана, в 3-5%
случаев синдром Ангельмана обусловлен однородительской дисомией материнского
происхождения и в 3-4 % – этот же синдром развивается вследствие мутаций
«импринтинга».
При обнаружении делеции или однородительской дисомии риск будет низким (около
1%), а при обнаружении мутаций импринтинга – значительным не только для
родителей, но и для ближайших родственников. В случаях «контрастных болезней» –
синдрома Беквита-Видемана и синдрома Сильвера-Рассела гены локализованы на
хромосоме 11 и центра импринтинга этой хромосомы.
Возможно существует связь геномного импринтинга с другими болезнями, в
частности с мультифакториальными болезнями (бронхиальная астма, атонический
дерматит, онкопатология, атопии и др.). Предполагают, что участие в подобных
патологических процесссах происходит специфическое метилирование цитозиновых
оснований ДНК, образующих антисмысловые транскрипты РНК, изменения нитронов и
др. Но скорее всего «виноваты» деструктивные дефекты генов в молекуле ДНК,
появляющиеся у потомка вследствие передачи ему пар хромосом только от отца или
только от матери. Если же передаются только половинки молекулы ДНК, то по другой
половинке возможно восстановление нарушенного гена (импритинг) и болезни не
возникает.
Все особи одного вида похожи друг на друга потому, что их биохимические циклы
схожи и вырабатывают одинаковые белки, из которых строятся одинаковые
биологические конструкции. Именно потому, что в пределах одного вида
биохимические циклы одинаковы, возможно половое размножение особей. Если
скрещиваются разные виды, то в геном потомка привносятся разнотипные генные
наборы, которые внесут разбаланс в тонкую настройку биохимических циклов плода.
В результате плод не сможет развиваться, если он возникнет в виде бластулы и
если вообще сперматозоид с его биохимией сможет проникнуть в яйцеклетку с другой
биохимией, чтобы было зачатие.
Недействующая же часть генома особей может отличаться от таковой у других
особей данного вида, причем чем больше, тем лучше. Но это никак не сказывается
ни на строении особей, ни на их способности к спариванию и производства
потомства, потому эта часть генома недействует (!) и не регулирует ни один
биохимический цикл.
Таким образом, есть множество способов размножения, но все они делятся на две
обширные группы:
- бесполое размножение (простое деление клеток),
- половое размножение (скрещивание) у растений и животных.
Самым древним способом является бесполое размножение. Оно происходит путем
репликации молекулы ДНК (повторение молекулы ДНК – построение её клона). Затем
для сохранения генной устойчивости молекулы ДНК при неблагоприятных условиях и
для увеличения генного наследия потомков (скрещивание двух разных одновидовых
половинок молекул ДНК для обогащения мусорной части генома потомка и для
исправления дефектов его действующей части) появилось половое размножение.
Бесполое размножение характерно для вирусов и простейших одноклеточных, хотя и у
бактерий можно наблюдать половой процесс (рис. 5).
Половое скрещивание характерно для многоклеточных, хотя даже у человека можно
наблюдать некоторые варианты бесполого размножения (однояйцевые близнецы). Из
этих механизмов только половое скрещивание может обеспечить увеличение генного
разнообразия геномов. Скрещивание возможно только внутри вида и особи передают
своему потомству не только настройки действующих наборов генов, но и
разнообразие генов недействующией («мусорной») части геномов.
Рис. 5. Виды размножения (слева) и бактериальный секс (справа). Пара
конъюгирующих бактерий, соединенных F-пилем – структурой, которую образует
клетка-донор.[21] Через F-пиль ДНК попадает в клетку-реципиент.
Недействующие части геномов у родителей могут содержать различающиеся наборы
генов. При половом скрещивании потомство получает «мусорные» гены со стороны
матери и со стороны отца и если эти наборы генов у матери и отца различны, то у
потомства получается более обогащенный геном. Это является основой появления
шанса отпочкования нового биологического вида от данного. Не зря половое
скрещивание называется скрещиванием – геномы матери и отца перекрещиваются,
обогащаясь при этом. Но обогащаются только недействующие части генома, потому
что в действующей части обогащаться нечему, так как материнская и отцовская
действующие части геномов по составу генов должны быть совершенно одинаковы. У
них могут быть разными только настройки генов. В действующей же части становится
возможным только исправление генетических дефектов, накопленных в филах матери и
отца.
Таким образом, именно генная избыточность, осуществляемая путем вирусного
заражения, является причиной появления новых видов живых систем. Если кто-то с
этим не согласен, то он должен доказать, что либо неверна центральная догма
молекулярной биологии, которая говорит о том, что информация передается только
от нуклеиновых кислот к белку, но не в обратном направлении,[7] либо
предложить какой-либо другой правдоподобный механизм появления новых генов в
геномах эволюционирующих живых систем, не противоречащий этой догме. А заодно и
объяснить, почему во всех геномах всех живых систем, начиная от самих вирусов и
вплоть до человека, присутствуют вирусные гены или их остатки. Теории мутагенеза
не проходят в силу выше рассмотренных противоречий.
Статья является одной из глав будущей книги «Законы природы и эволюция
Вселенной»
Список литературы
- Acevedo, Oscar L.; Andrews, Robert S. Synthesis of propane-2,3-diol
combinatorial monomers. Tetrahedron Letters (1996), 37(23), 3931-3934.
- Bernard La Scola et al. The virophage as a unique parasite of the giant
mimivirus // Nature. 2008. V. 455. P. 100–104.
- Brown, S. W.; U. Nur (1964). «Heterochromatic chromosomes in the coccids».
Science 145: 130–136.
- Cairns-Smith, A.G. Towards a Theoretical Biology / Waddington, C,H.. —
Edinburgh University Press, 1968. — Vol. 1. — P. 57–66.
- Carl Woese. The Genetic Code. — Harper & Row, 1968.
- Chetverin, A.B., Chetverina, H.V., Munishkin, A.V. (1991) J. Mol. Biol.,
222, 3–9.
- Crick, F.H.C. (1958): On Protein Synthesis. Symp. Soc. Exp. Biol. XII,
139—163.
- Crick, F. (1970): Central Dogma of Molecular Biology. Nature 227, 561—563.
- Dawkins Richard Origins and miracles // The Blind Watchmaker. — New York:
W. W. Norton & Company, 1996.
- DeChiara, Thomas M.; Elizabeth J. Robertson and Argiris Efstratiadis
(February 1991). «Parental imprinting of the mouse insulin-like growth factor
II gene». Cell 64 (4): 849–59.
- Defne Arslan, Matthieu Legendre, Virginie Seltzer, Chantal Abergel, and
Jean-Michel Claverie. Distant Mimivirus relative with a larger genome
highlights the fundamental features of Megaviridae. NAS 2011 108 (42)
17486-17491.
- Fischer MG1, Suttle CA. A virophage at the origin of large DNA transposons.
Science. 2011 Apr 8;332(6026):231-4.
- Fisher R. A. The genetical theory of natural selection. — 2-nd ed. — New
York, 1958.
- Frederick R. Blattner*, Guy Plunkett III*, Craig A. Bloch, Nicole T. Perna,
Valerie Burland, Monica Riley, Julio Collado-Vides, Jeremy D. Glasner,
Christopher K. Rode, George F. Mayhew, Jason Gregor, Nelson Wayne Davis,
Heather A. Kirkpatrick, Michael A. Goeden, Debra J. Rose, Bob Mau and Ying
Shao. The Complete Genome Sequence of Escherichia coli K-12. Science 5
September 1997: Vol. 277 no. 5331 pp. 1453-1462.
- Fuhrman J. A. (1999) Marine viruses and their biogeochemical and
ecological effects. Nature 399: 23. 541–548.
- Gilbert, Walter (Feb 1986). "The RNA World". Nature 319: 618.
- Goldschmidt R. The material basis of evolution. New Haven: Yale Univ.
Press, 1982. (2nd d.).
- Hughes-Schrader, S. (1948). «Cytology of coccids (Coccoïdea-Homoptera)».
Advances in Genetics 35 (2): 127–203.
- Jack W. Szostak. Origins of life: Systems chemistry on early Earth //
Nature. 2009. V. 459. P. 171–172.
- La Scola B1, Desnues C, Pagnier I, Robert C, Barrassi L, Fournous G,
Merchat M, Suzan-Monti M, Forterre P, Koonin E, Raoult D. The virophage as a
unique parasite of the giant mimivirus. Nature. 2008 Sep 4;455(7209):100-4.
- Matxalen Llosa, F. Xavier Gomis-Rüth, Miquel Coll2 and Fernando de la
Cruz. Bacterial conjugation: a two-step mechanism for DNA transport (micro
review). Molecular Microbiology (2002) 45(1), 1–8.
- Matthew W. Powner, Béatrice Gerland, John D. Sutherland. Synthesis of
activated pyrimidine ribonucleotides in prebiotically plausible conditions //
Nature. 2009. V. 459. P. 239–242.
- Morgan Gaia, Philippe Colson, Christelle Desnues, Bernard La Scola. The
Virophage Concept. Wiley Online Library, Copyright © 2001 John Wiley & Sons,
Ltd. Published Online: 15 MAY 2013.
- Nelson, K.E., Levy, M., and Miller, S.L. Peptide nucleic acids rather than
RNA may have been the first genetic molecule (2000) Proc. Natl. Acad. Sci. USA
97, 3868–3871.
- Nur, U. (1990). «Heterochromatization and euchromatization of whole
genomes in scale insects (Coccoidea: Homoptera)». Dev. Suppl.: 29–34.
- Schoning, K; Scholz P; Guntha S; Wu X; Krishnamurthy R; Eschenmoser A
(November 2000). "Chemical etiology of nucleic acid structure: the alpha-threofuranosyl-(3'->2')
oligonucleotide system.". Science 290 (5495): 1347.
- Seita, Tooru; Yamauchi, Kiyoshi; Kinoshita, Masayoshi; Imoto, Minoru.
Condensation polymerization of nucleotide analogues. Die Makromolekulare
Chemie (1972), 154:255-261.
- Vladimir A. Belyi, Arnold J. Levine, Anna Marie Skalka. Unexpected
Inheritance: Multiple Integrations of Ancient Bornavirus and Ebolavirus/Marburgvirus
Sequences in Vertebrate Genomes. PLoS Pathogens. July 2010, Volume 6, Issue 7.
- Weinbauer M. G. (2004) Ecology of prokaryotic viruses. FEMS Microbiol.
Rev. 28: 127–181.
- Wilkinson, Lawrence S.; William Davies and Anthony R. Isles (November
2007). «Genomic imprinting effects on brain development and function». Nature
Reviews Neuroscience 8 (11): 832–843.
- Wilson W. H., Mann N. H. (1997) Lisogenic and lytic viral production in
marine microbial 49. communities. Aquat. Microb. Ecol. 13(1): 95–100.
- Wolfsberg T, McEntyre J, Schuler G (2001). «Guide to the draft human
genome». Nature 409 (6822): 824 – 6.
- Аршинов В. И. Синергетика как феномен постнеклассической науки. — М.,
1999.
- Бернал Дж. Возникновение жизни, М., 1969.
- Воронцов Н. Н. Синтетическая теория эволюции: её источники, основные
постулаты и нерешенные проблемы // Журн. Всес. хим. о-ва им. Д. И. Менделеева.
1980. Т. 25. N 3. С. 293-312.
- Гайдес М.А. Общая теория систем (Системный анализ). Глобус-пресс. 2004.
- Гродницкий Д. Л. Две теории биологической эволюции. — Саратов, 2002.
- Копылов А.И. Роль вирусов в структуре и функционировании микробных
сообществ в пресноводных экосистемах. Journal of Siberian Federal University.
Biology 4 (2013 6) 354-367.
- Костецкий ЭЛ. О возможности синтеза нуклеопротеинов на матрице апатита /
Э.Я. Костецкий, С.А. Алексаков // Докл. АН СССР. 1981. Т. 260. С. 1013-1018.
- Хакен Г. Синергетика. — М.: Мир, 1980. — 406 с.
- Четверин А.Б. // Успехи биол. химии. 1998. T.38. С.3—75.
- Четверина Е. В., Четверин А. Б.. НАНОКОЛОНИИ: ОБНАРУЖЕНИЕ, КЛОНИРОВАНИЕ И
АНАЛИЗ ИНДИВИДУАЛЬНЫХ МОЛЕКУЛ. Успехи биологической химии, т. 48, 2008, с.
3–643.
Если вы заметили орфографическую, стилистическую или другую ошибку на этой странице, просто выделите ошибку мышью и нажмите Ctrl+Enter. Выделенный текст будет немедленно отослан редактору
|
|
|
|