Данная информация предназначена для специалистов в области здравоохранения и фармацевтики. Пациенты не должны использовать эту информацию в качестве медицинских советов или рекомендаций.
Глава 3. Метаболические миры
§1. Мир растений
1) Простой соматический и простой вегетативный БУС (простые соматический
и вегетативный автоматы).
Растение – это составная (двойная) многофункциональная система, состоящая из:
того же простого автомата мира минералов с теми же функциями и
системы обмена веществ и ее собственного управления, которая постоянно
определяет разрушения СФЕ системы и стимулирует действия СФЕ системы обмена
веществ для восстановления разрушенных структур (рис. 33).
Чтобы не путаться в названиях простой БУС растений назовем простым
соматическим БУС, а простой БУС, управляющий системой (подсистемой)
обмена веществ – простым вегетативным БУС. Итого, у растений есть
два БУС – соматический и вегетативный. Соматический БУС «отвечает» за управление
структур растения (волокна, ткани, полости и пр.), определяющих
физическое состояние растения – его форму, гибкость и эластичность и пр.
Вегетативный БУС следит за целостностью этих структур растения.
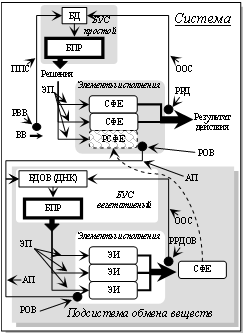
Рис. 33. Блок-схема растения как системы с простым БУС и
управлением системы (подсистемы) обмена веществ.
ВВ – внешнее воздействие; РСФЕ – разрушенная СФЕ (темного цвета); ЭИ –
элементы исполнения (РНК и клеточные органеллы); АП – афферентные пути; ЭП –
эфферентные пути; РОВ – рецепторы обмена веществ; РРДОВ – рецепторы результата
действия обмена веществ; БДОВ – «база данных обмена веществ» (ДНК). Пунктирная
линия – новая СФЕ вырастает на месте разрушенной.
Соматический БУС выполняет ту же функцию управления, что и простой БУС у
минералов – точно так же управляет исполнительными элементами. Но система обмена
веществ, входящая в состав растений, обеспечивает сохранность его элементов
исполнения, постоянно восстанавливая разрушенное. Растения отличаются от систем
минерального мира только наличием системы обмена веществ, основанной на генном
механизме регенерации и репликации и этот механизм имеет собственный БУС,
управляющий системой обмена веществ.
Основной закон систем мира растений – закон замены разрушенных частей,
научить Я систем менять собственный состав для самовосстановления после
разрушений и самовоспроизведения, используя для этого обмен веществ.
Растения – это как бы две системы в рамках одной. Одна из них является
обычной системой с простым БУС (соматическим БУС), а другая является системой с
подобным же БУС, но у которого есть отличия, дающие право назвать этот БУС
вегетативным. Его рецепторами внешнего воздействия (РВВ) являются РОВ, внешним
воздействием для него являются изменения внутренней реальности системы, а
результатом действий системы обмена веществ – восстанавливаемая СФЕ
(регенерируемая). Так как система обмена веществ является неотъемлемой частью
самой системы, то элементы исполнения системы обмена веществ также входят в
понятие «внутренняя реальность». Это значит, что вегетативный БУС с помощью РОВ
получает информацию и о собственных элементах, т.е., о своей внутренней
реальности, и знает ее, если может определить дефекты и «починить» их.
Управление системой основного обмена основано на оценке внутренней реальности,
т.е., на ее осознании. Осознание внутренней реальности является
предсознанием. Следовательно, растения имеют два простых БУС
(соматический и вегетативный), из которых один из них обладает предсознанием
(вегетативный).
Алгоритмы соматического и вегетативного БУС (рис. 34).
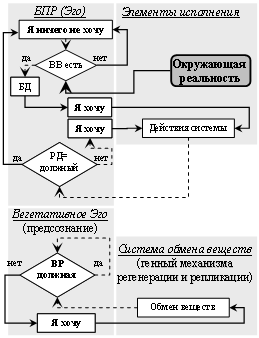
Рис. 34. Алгоритм работы БПР простого и вегетативного БУС у
систем мира растений и их взаимодействие.
БД – «база данных»; РД=БД – результат действия равен заданному и
описанному в «базе данных»; ВР должная – внутренняя реальность должная;
восстановление ВР – восстановление внутренней реальности.
Алгоритм соматического БУС точно такой же, как и алгоритм БПР простого БУС у
систем минерального мира. Если нет разрушений СФЕ, то в его алгоритме нет
никаких отличий. Отличия возникают, когда происходит разрушение каких-либо СФЕ.
Тогда цикл действий «РД=должный – хочу действия системы» будет продолжаться
дольше, потому что будет не хватать СФЕ для быстрого достижения цели.
Но параллельно в системе обмена веществ будет происходить другой цикл действий –
«ВР=должная – хочу обмен веществ» до тех пор, пока разрушенные СФЕ не будут
восстановлены. После восстановления разрушенных СФЕ отличия в алгоритме опять
исчезают. Фактически это параллельная работа двух Эго – соматического и
вегетативного Эго (предсознания). Они работают относительно независимо друг от
друга, но в кооперации: соматический БУС использует СФЕ системы для своих
«нужд», а вегетативное Эго следит за сохранностью этих СФЕ, восстанавливая их по
мере надобности.
«Должная ВР» вегетативного БУС дает информацию о том, какой должна быть
внутренняя реальность. В «базе данных внутренней реальности» (ДНК) есть описание
параметров внутренней реальности, которые должны быть у нормальной системы.
Наличие этой базы дает возможность сравнения параметров актуальной внутренней
реальности с должной, т.е., оценки внутренней реальности. А оценка какой-либо
реальности является частью сознания. У простого (соматического) БУС растений нет
осознания обеих реальностей, поэтому у него нет и сознания. Но у вегетативного
БУС есть осознание только внутренней реальности, но нет осознания окружающей
реальности. Поэтому это осознание внутренней реальности является
предсознанием.
Элементами исполнения системы обмена веществ являются все виды РНК с
рибосомами и другие различные внутриклеточные органоиды, выполняющие функции
трансмембранной диффузии и внутриклеточного пищеварения. РНК снимают информацию
с генов ДНК и с помощью сложного механизма рибосом синтезируют белки для
последующей постройки из них элементов разрушенных СФЕ. А внутриклеточные
органиоиды осуществляют поставку необходимых веществ и удаление расходного
материала из клетки за счет систем внутриклеточного пищеварения.
В остальном системы с простым БУС с управлением обмена веществ также, как и
системы минерального мира, «ощущают» внешнее воздействие, измеряют
его и в зависимости от величины этого воздействия вырабатывают столько
результата действия, сколько задано отношением вход-выход [8, 9].
Параметры этого задания так же находятся в «базе данных». Система мира растений
может вырабатывать столько результата действия, сколько нужно (оптимально), от
минимума до максимума, в зависимости от задания, и тоже с определенной
точностью, потому что у нее есть отрицательная обратная связь (ООС), которая
постоянно измеряет результат действия, сравнивает его с заданным и если есть
отличие от заданного, так воздействует на исполнительные элементы, чтобы
различие исчезло. Задание так же находится в «базе данных» и оно внедряется туда
в момент «рождения» системы.
Назначением этой схемы было показать только те факторы, которые прямо влияют
на «хочу-не-хочу» БПР, поэтому данная схема алгоритма не детальная и здесь нет
описания всех действий БПР для продолжения действий системы до равенства
актуального результата действия заданному.
Его выбор между хочу и не хочу все еще во многом определен внешними
критериями, но часть его решений основана уже на его собственных действиях – он
определяет состояние внутренней реальности и на основе этих определений сам
решает, какие действия он должен сделать для восстановления разрушенных
структур. Он уже не так подвержен разрушающему влиянию внешней среды, потому что
он может восстанавливать разрушенное. Следовательно, вегетативный БПР простого
БУС еще в большей степени автономен.
В этом главное отличие алгоритма вегетативного БУС от алгоритмов предыдущих
БУС. Этот БУС постоянно контролирует внутреннюю реальность систем и
восстанавливает ее, в случае разрушения. Для этого используется генный механизм
регенерации системы обмена веществ.
Следует отметить, что система обмена веществ растений и животных отличается
тем, что у растений есть только внутриклеточный генный механизма регенерации
и репликации, а у животных, кроме внутриклеточного генного механизма
регенерации и репликации есть еще и многоклеточные внутренние органы.
Поэтому у растений управлением системы обмена веществ занимается внутриклеточный
простой БУС (ДНК, РНК и рибосомы), а у животных этим управлением занимается
отдельная специальная система – сложный вегетативный БУС со своими инстинктами
(вегетативная нервная система).
Простой соматический БУС (простой соматический автомат мира
растений).
Растения как физическое тело ничем не отличаются от систем минерального мира,
простые автоматы мира растений точно такие же, как и простые автоматы мира
минералов. То же касается и соматического БУС растений, он такой же как и
простой БУС систем мира минералов, который мы уже рассмотрели в предыдущей
главе. Отличие растений от систем мира минералов только в наличии у них системы
обмена веществ, которая может обеспечить размножение растений и восстановление
их разрушенных структур. Другими словами, растения – это минералы, которые
научились размножаться и восстанавливаться. Это стало возможным потому, что у
растений появилась система (подсистема) обмена веществ, которая имеет простой
вегетативный БУС.
Реакция системы с простым соматическим БУС – это та же реакция простого
автомата мира минералов с той же целью – выработка определенного по количеству и
качеству целевого результата действий с заданной точностью. Простой соматический
автомат реализует то же право на точное действие по закону – «сколько
нужно, столько будет сделано», с точностью до одного кванта действия его СФЕ.
Простой вегетативный БУС (простой вегетативный автомат мира
растений).
Реакция простого вегетативного автомата мира растений направлена на изменения
не в окружающей, а во внутренней реальности. Его цель – восстанавливать
разрушенные структуры системы. Простой вегетативный автомат реализует право на
точное действие внутри системы по принципу: «каждая разрушенная
структура должна быть восстановлена». Но простой вегетативный автомат – это
тот же простой автомат мира минералов, только с другой точкой приложения.
Поэтому реализация этого права осуществляется по тому же закону простого
автомата – «сколько нужно, столько будет сделано», с точностью до одного
кванта действия его СФЕ.
Цель достигается счет восприятия внутренней реальности, ее оценки и работы
системы (подсистемы) обмена веществ, восстанавливающей разрушенные элементы
исполнения систем.
Абсолютно все объекты Вселенной подвержены разрушающему влиянию окружающей
реальности. Поэтому, все, что было сказано в отношении устойчивости систем с
простым БУС к разрушающему действию окружающей реальности, касается всех
остальных систем в той же мере. И если системы мира минералов (с простым БУС)
«изобрели» только пассивные методы защиты, то системы мира растений (с таким же
простым БУС, но имеющим в добавок систему обмена веществ и ее управление) имеют
уже активные способы защиты – они могут восстанавливать разрушенные СФЕ.
Мы не будем рассматривать все частности генного механизма системы обмена
веществ, для этого есть достаточно описаний в литературе [36], потому
что нашей задачей является только показ особенностей блоков управления систем.
Заметим лишь, что исходя из этих особенностей необходимо искать в генном
механизме регенерации и репликации те элементы, которые необходимы для работы
такого БУС. Это касается рецепторов (РОВ), которые определяют внутриклеточные
разрушения (нарушения внутренней реальности), и той «базы данных», в которой
находится описание самих структур внутренней реальности. Эти вопросы в отношении
внутриклеточной системы обмена веществ в научных исследованиях пока еще вообще
не были затронуты.
Состав простого вегетативного БУС.
Состав элементов управления систем включает в себя:
- «базу данных обмена веществ» (БДОВ),
- блок принятия решений (БПР),
- ОСС,
- рецепторы результата действия обмена веществ (РРДОВ),
- рецепторы обмена веществ (РОВ) и
- пути информации с РОВ (афферентные пути, АП).
В состав растений, в отличие от минералов, добавилась подсистема (система)
обмена веществ с ее собственным вегетативным БУС, обладающим предсознанием, и
его элементами – РОВ и АП и это превратило систему с пассивной
защитой от разрушения в систему с активной защитой от разрушения.
Системы научились самовосставливаться.
Эволюционная роль вегетативного БУС.
Роль вегетативного БУС в эволюции сознания в том, что впервые появилось Я,
у которого есть предсознание – осознание внутренней реальности системы.
Роль систем с вегетативным БУС (растений) в эволюции Вселенной в том, что
в ней появились объекты, которые могут восстанавливать свои разрушенные
элементы и реплицировать самих себя. Этим растения заложили основу появления
остальных миров живых систем – животных и человека.
Вегетативный БУС отличается от предыдущих тем, что он «осознает» внутреннюю
реальность системы (свою внутреннюю реальность). У растений появилось
предсознание. Растения являются очередным этапом эволюции от простых
систем к человеку и появление в них предсознания дало возможность в дальнейшем,
у животных, появлению и развитию сознания. Кроме того, если минералы создали
базу – построили планеты со всем необходимым для жизни содержимым и выработали
все необходимые элементы для появления жизни (РНК, органику), на которой смогли
появиться растения, то растения создали кислородную атмосферу, в которой смогли
появиться и развиваться животные, и дали энергию и материалы (высококалорийную
пищу) животным. Другими словами, растения подготовили условия на Земле для
появления животных, дав кислородную атмосферу и калорийную пищу.
Растения появились не сразу. Если на создание первых систем мира
минералов с простым БУС (атомов гелия и водорода) ушло не более минуты,
то на создание первых систем мира растений ушло более 15 млрд лет.
Столько времени ушло, чтобы простой БУС «научился» воспринимать и осознавать
внутреннюю реальность системы и управлять ею.
Первыми растениями были одноклеточные водоросли. Но это уже были полноценные
растения с полным набором всех элементов, необходимых для регенерации клеточных
элементов и репродукции (размножения) самих клеток.
Мы не будем здесь рассматривать прокариотов и эукариотов, наша цель несколько
иная – показать сложность растительной клетки и ее отличия от любых других тел и
объектов минерального мира. Основное строение (анатомия) клеток включает в себя
наружную мембрану, обеспечивающую постоянство внутреннего состава
клетки путем избирательной диффузии в обоих направлениях через нее, внутреннюю
строму клетки, обеспечивающую поддержание определенной формы,
органеллы клетки, обеспечивающие автотрофию (клеточное пищеварение) и
генный механизм регенерации и репродукции. Физически клетка
представляет собой каплю (кусочек) сложного структурированного геля,
ограниченного от окружающей реальности мембраной. Химический состав этого геля –
80-90% воды, остальное – нуклеотиды (ДНК и РНК), белки, липиды, углеводы и
минеральные соли. ДНК и РНК вместе с рибосомами участвуют в синтезе белка. Белки
составляют строму клетки и ее энзимы. Жиры являются одним из подклассов липидов,
входят в состав наружной и внутренних мембран, а углеводы также входят в состав
стромы, используются в качестве источника энергии и образуются в растениях в том
числе путем фотосинтеза. Минералы участвуют в поддержании осмотических давлений
в клетке и играют роль переносчиков информации (внутриклеточная электрохимия)
механизма возбуждения клетки.
Большинство этих составных частей клетки, включая аминокислоты, можно найти в
природе отдельно от клетки, даже в межпланетной и межзвездной пыли. Это говорит
о том, что элементы растительной клетки появились в виде минеральных систем еще
задолго до появления самой клетки. Даже сама клеточная форма, капли геля
(коацерваты), появились задолго до появления самих растительных клеток.
Следует обратить на весьма существенное обстоятельство. Вероятно, все знают,
что практически все живые системы, включая наше тело, содержат очень много воды.
И, вероятно, многие думают, что это наш минус. Вот будь мы из железа, было бы
лучше. Но это не так. Большинство реакций органической и биохимии могут
осуществляться только в растворах, и из растворных реакций подавляющее
большинство могут проходить только в водных растворах. Следовательно, для
нормального функционирования внутриклеточной деятельности клетка должна быть
заполнена водой. Поэтому практически все живые существа в своем составе содержат
очень много воды и этим решается очень много проблем. Все твердые тела очень
трудно меняют форму и сделать из них новую СФЕ с целью замены разрушенной СФЕ
очень трудно и энергетически неоправданно. Из водяного пузыря сделать что-то
можно без проблем. Вода мягкая и не «ломается» при ударах. Поэтому, в условиях
агрессивной окружающей реальности иметь «водяное» тело выгодней, чем «железное».
Да, при некоторых воздействиях легко возникают разрушения, но они также легко
возмещаются теми же водными формами. Короче, слепить скульптуру из воды намного
легче, чем из кристалла. Да, зато кристалл легче держит свою форму, чем вода,
которая растекается. Но если воду заключить в мембрану, то и форму можно
сохранить, и к ударам вода устойчивее, чем кристалл, который при ударах
ломается, и новую форму легче сделать.
Еще до появления клеток все органические и начала биохимических реакций
происходили в океанах, в водной среде. В последующем, когда клетки отграничились
от окружающей реальности мембраной, они включили в себя как бы капельку того
океана, в котором они когда-то появились. Поэтому у большинства живых существ
минеральный состав клеточной плазмы и плазмы крови очень напоминает минеральный
состав первобытных морей. Отсюда, жизнь могла возникнуть только в океанах.
Концепцию коацерватов впервые предложил А.И.Опарин, выступив 3 мая 1924 года
на собрании Русского ботанического общества с докладом «О возникновении жизни»,
в котором предложил теорию возникновения жизни из бульона органических веществ.
В качестве протоклеток он рассматривал коацерваты — органические структуры,
окружённые жировыми мембранами.
Коацервация (от лат. coacervatio — собирание, накапливание) —
расслоение коллоидной системы с образованием коллоидных скоплений (коацерватов)
в виде двух жидких слоев или капель. Коацервация может возникать в результате
частичной дегидратации дисперсной фазы коллоида, являясь начальной стадией
коагуляции [60].
Сущность явления коацервации заключается в отмеривании из однородного
коллоидного раствора слоя или капель, связанном с переходом от полного
смешивания к ограниченной растворимости.
Растения умеют восстанавливать разрушенные структуры. Чтобы восстановить
разрушенное нужно знать, как и из чего построить
вновь это разрушенное. Для этого необходимы три компоненты:
знание данных того, что нужно построить,
инструменты, с помощью которых можно это строить и
исходные материалы, из которых это будет построено.
В растительных клетках все эти компоненты есть. Ими являются:
знание данных – «базы данных», в которых хранится информация (данные) о
строении всех структур клетки (ДНК),
инструменты – различные виды РНК, матричные, рибосомные и транпортные,
плюс большое множество «малых РНК» – все это своеобразные молекулярные
инструменты, которые снимают информацию с ДНК, способны к самовоспроизведению
и переносу исходных материалов в нужное место и каталитическому синтезу
клеточных белков,
исходные материалы – поставляет аппарат внутриклеточного пищеварения
(энзимы и органеллы вместе с трансмембранной диффузией).
Все необходимые исходные низкомолекулярные материалы, из которых
синтезируются высокомолекулярные белки, поставляются извне за счет их
переноса свободным притоком жидкости к растению, всасывания
их растением за счет диффузии через мембраны и внутриклеточного синтеза.
Это и есть трехкомпонентный высоконадежный и точный генный механизм
молекулярной регенерации и репликации клеточных структур. Основой этого
механизма является способность некоторых его элементов к молекулярному
самовоспроизведению. Белки являются одним из главных компонентов любой живой
клетки, но они чрезвычайно неустойчивы во внешней среде. Для постоянного
строительства белковых молекул и был создан генный механизм регенерации и
репликации.
Нет сомнения, что мир растений произошел из мира минералов, но не совсем
ясно, каким образом возникли первые устойчивые самовоспроизводящиеся системы. В
живых организмах практически все процессы происходят в основном благодаря
энзимам белковой природы. Белки, однако, не могут самореплицироваться и
синтезируются в клетке de novo только на основании информации, заложенной в ДНК.
Но и удвоение ДНК происходит только благодаря участию белков и РНК. Образуется
замкнутый порочный круг, из-за которого, в рамках теории
самозарождения жизни необходимо признать не только абиогенный синтез обоих
классов молекул (белков и ДНК), но и спонтанного возникновения сложной системы
их взаимосвязи, что крайне маловероятно.
Очевидно, сначала должен был быть механизм, поднявший химические и
физико-химические интеракции систем мира минералов до стойкого и
самодостаточного уровня органической и биохимии систем без участия ДНК. Эволюция
систем от мира минералов до уровня мира растений – это химическая
эволюция, возникновение и развитие тех молекул, которые имеют значение в
историческом развитии живых существ. При этом простые органические,
пребиотические молекулы возникли из неорганических молекул под влиянием
энергетических процессов и информационного шума (шумового «дребезга», см. выше).
И шумовой «дребезг» является неотъемлемым атрибутом работы любой системы, что
обусловлено особенностями строения и функции систем – дискретностью (квантами)
их результатов действия. Следовательно, сложные органические молекулы получились
из простых органических химических соединений за счет притока энергии извне и
«врожденных» свойств этих соединений (они были такими созданы), позволяющих
«собирать» из них высокомолекулярные соединения. А врожденные свойства – это и
есть дискретность результата действия систем.
В качестве источников энергии рассматриваются ультрафиолетовое излучение
Солнца, тепло вулканических процессов, ионизирующее излучение радиоактивного
распада и электрические разряды (молнии). Есть надежные эмпирические и
экспериментальные подтверждения этих гипотез, поскольку органические молекулы
обнаружены даже в межзвездной пыли [45, 53].
Предполагалось (Опарин А.И., 1932, [29]), что эта органика
получалась в первичном «бульоне» первобытных океанов Земли под действием
вулканического тепла и энергии электрических разрядов (молний, рис. 35) .
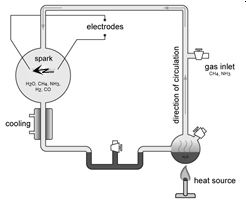
Рис. 35. Эксперимент Миллера-Юри (1953) [14].
Известно, что многие органические молекулы, включая аминокислоты (составные
элементы белков), могли формироваться из неорганических соединений в условиях
древней Земли (эксперимент Миллера-Юри, 1953).
Из появляющихся в условиях этого эксперимента промежуточных продуктов
альдегидов и синильной кислоты HCN можно получить при симулируемых условиях
земли 4,5 млрд. лет назад дальнейшие биомолекулы. Таким образом Хуан Оро удалось
в 1961 получить синтез аденина:
| Эдукты |
|
|
Продукты |
| 2 Формальдегид |
HCN |
H2O |
Серин |
| 5 Формальдегид |
|
|
Рибоза |
| |
5 HCN |
|
Аденин |
Из рибозы, аденина и трифосфата возникает аденозинтрифосфат (АТФ), который
используется в организмах в качестве универсального энергоносителя и
строительного элемента (как монофосфат) рибонуклеиновых кислот (РНК).
Существуют и альтернативные гипотезы, например, теория Вехтерхойзера
[58]. Эта теория хорошо согласуется с известными фактами жизни бактерий в
условиях глубоководных термальных источников (черные курильщики), именно тех
условий, которые были на молодой Земле. Древнейшие из сегодня живущих
микроорганизмов самые жароустойчивые, предельный известный температурный
максимум для их роста составляет +113 °C. Кроме того железо-серные активные
центры и сегодня задействованы в биохимических процессах, что может указывать на
первичное участие Fe-S-минералов в развитии жизни (рис. 36).
Таким образом, вопрос об источнике энергии для полимеризации макромолекул
решается относительно просто, таких источников могло быть несколько. Но для
самоподдержания жизни одних источников энергии недостаточно. Еще до появления
первых клеток низкомолекулярные соединения должны были каким-то образом
полимеризоваться до высокомолекулярных соединений, вплоть до белков, чтобы
построить из этих соединений различные структуры. Кроме того они должны уметь
размножаться, иначе каждая вновь созданная структура рано или поздно разрушалась
бы и исчезала. Здесь вообще нет речи о восстановлении разрушенных
внутриклеточных структур, только первичный синтез и примитивное размножение
первичных молекулярных агрегатов и ансамблей.
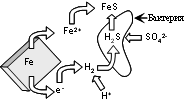
Рис. 36. Железо-серная хемоавтотрофия.
Источник энергии из восстановления частично окисленных железно-серных
минералов, например пирита (FeS2), водородом (уравнение реакции: FeS2 + H2 FeS +
H2S) для эндотермического синтеза мономерных структурных элементов биомолекул и
их полимеризации [14].
Есть множество теорий, каким образом могли образоваться высокомолекулярные
образования в условиях ранней Земли. Показано, что:
Кристаллические поверхности могут служить матрицей для растущих
макромолекул. При этом различные кристаллические поверхности могут связывать
определенные энантиомеры молекул. L-и D аминокислоты присоединяются к разным
местам кристалла кальцита.
Аарон Качальсский (Вайцманн-институт, Израиль) показал, что в водном
растворе, содержащем монтмориллонит (один из минералов глин), возможен синтез
белков с длиной цепи в больше чем 50 аминокислот с почти 100%-ным выходом.
Ионы металлов могут выступать в роли катализаторов, доноров электронов или
же включаться в биомолекулы.
Минералы глин в водных растворах часто несут поверхностный электрический
заряд и могут таким образом притягивать и удерживать противоположно заряженные
органические молекулы.
В микрополостях горных пород молекулы органических соединений защищены от
ультрафиолетового облучения.
Может быть когда-то и где-то это сработало и появились первые молекулы РНК.
Но сейчас, вероятно, все эти модели для синтеза белков сегодняшними живыми
клетками почти не используются, а вместо них используется генный механизм
регенерации и репликации, использующий ДНК. Следовательно, первые организмы
должны были развить этот механизм еще до появления ДНК, потому что сомнительно,
чтобы у первых организмов уже была ДНК. Во-первых, в ДНК
«записана» очень обширная информация, накопленная за длительный период эволюции
живых клеток и этот период требовалось сначала прожить. Во-вторых,
генный механизм с использованием ДНК слишком сложный и многоступенчатый, чтобы
он появился с самого начала. ДНК нужно было сначала построить и только затем
эксплуатировать.
Иначе вопрос обстоит в отношении молекул РНК, которые способны к хранению,
передаче, и размножению генетической информации. В начале 1980-х в лаборатории
Т. Чека и С. Олтмана в США была открыта каталитическая способность РНК. По
аналогии с энзимами (ферментами) РНК-катализаторы были названы рибозимами,
за их открытие была присуждена Нобелевская премия 1989. Также РНК способны
создавать двойную цепочку и самореплицироваться.
Некоторые из них имеют способность катализировать синтез белков, которые, в
свою очередь, сами катализируют синтез РНК и свой собственный синтез (развитие
трансляции). Некоторые молекулы РНК соединяются в двойную РНК-спираль, они
развиваются в молекулы ДНК и носители наследственной информации (развитие
транскрипции). Рибосомы считаются первоначальными клеточными органоидами, это
открытие считается важным взносом в обоснование гипотезы мира-РНК. Уже можно
уверенно сказать, что молекулы РНК, по меньшей мере принципиально, в состоянии
синтезировать белки из аминокислот.
Необходимо было чрезвычайное стечение обстоятельств, чтобы в одной
коацерватной капле собрались все факторы и минимум всех необходимых элементов,
чтобы получилась первичная самая простая и примитивная растительная клетка,
способная к автотрофии и генной репликации. Но не зря на ее создание
потребовалось около 15 млрд лет эволюции. По закону больших чисел то, что в
принципе могло произойти, за такой длительный срок произошло и появилась первая
живая клетка. Очень вероятно, что первые клетки вообше не умели регенерировать
свои разрушенные структуры, потому что это требует уже достаточно сложного
механизма поиска необходимых участков на РНК или ДНК для съема информации и его
управления (сложный вегетативный БУС). Но они могли реплицировать самих себя
(размножаться), используя ту же информацию с тех же носителей (с РНК),
считываемую всю сразу за один проход. И только потом, когда уже саморазмножаемые
автотрофные клетки развились, их эволюция привела к появлению более тонких
механизмов генной регенерации, позволяющих определять внутриклеточные разрушения
и регенерировать разрушенные структуры путем считывания только выборочной части
необходимой генетической информации, отыскивая для этого специфические участки
ДНК.
Таким образом, РНК могли существовать полностью автономно, катализируя
«метаболические» реакции, например, синтеза новых рибонуклеотидов и
самовоспроизводясь, сохраняя из «поколения» в «поколение» каталитические
свойства. Информационный шум привел к накоплению случайных мутаций и появлению
РНК, катализирующих синтез определённых белков, являющихся более эффективным
катализатором, в связи с чем эти мутации закреплялись в ходе естественного
отбора. С другой стороны возникли специализированные хранилища генетической
информации — ДНК. РНК сохранилась между ними как посредник.
Реликты РНК мира остались и в современных клеточных носителях энергии (АТФ —
рибонуклеотид, а не дезоксирибонуклеотид) и в путях синтеза самих
дезоксирибонуклеотидов: им всегда предшествуют рибонуклеотиды, для удвоения ДНК
необходимы РНК-«затравки».
Все это дало пищу для предположений существования сначала РНК-мира, после
которого, в результате его эволюции и появился современный мир ДНК. Вполне
вероятно, что РНК-мир был вершиной развития мира минералов, после которого
появился ДНК-мир и основанный на нем мир растений.
О том как выглядели самовоспроизводящиеся РНК-системы есть разные
предположения. Чаще всего постулируется необходимость агрегирующих РНК мембран
или размещения РНК на поверхности минералов и в порах рыхлых пород. В 1990-х А.
Б. Четвериным с сотрудниками была показана способность РНК формировать
молекулярные колонии на гелях и твёрдых субстратах при создании им
условий для репликации. Происходил свободный обмен молекулами, которые при
столкновении могли обмениваться участками, что показано экспериментально. Вся
совокупность колоний в связи с этим быстро эволюционировала.
РНК молекулы в колониях на гелях были подобны живым, но не были живыми. Они
были всего лишь вершиной эволюции мира минералов – органикой, способной
размножаться. Эта группа химических элементов не зря называется органикой, из
них, в дальшейшем, будут построены организмы всех живых систем. Органический
материал был создан еще до того, как из него были построены организмы.
Следовательно, мир растений начался в мире минералов еще до того, как
появились сами растения. И это также указывает еще на то, что изначально
истинные элементарные частицы минерального мира были сделаны с такими свойствами
и для их действий им были заданы такие законы природы, что эволюция мира
минералов однозначно пришла к появлению РНК-мира. Это все является не
самоорганизацией, и самосборкой по заданной «дорожке»,
т.е., принудительной извне организации. Мир растений просто обязан был появиться
в результате эволюции мира минералов, потому что он был таким сделан.
Следовательно, появление мира растений было заранее предопределено всем
развитием мира минералов.
Колонии молекул РНК, «поселившиеся» в коацерватных каплях и умеющие создавать
белки и ферменты, развивались успешнее. Лучше выживали те, которые могли
синтезировать лучшие комбинации ферментов, тем самым начался естественный отбор,
закрепляемый в генах сначала РНК, а затем и в более надёжном механизме хранения
информации – в ДНК. И, наконец, отделившиеся от внешнего мира фосфолипидной
мембраной, препятствующей рассеиванию своих молекул, получилась первичная
примитивная клетка, пока еще почти лишенная органоидов, но в протоплазме которой
уже был необходимый («джентльменский») набор ферментов и содержащая генный
механизм репликации, возможно вначале основанный только на РНК, но в дальнейшем
дополненный ДНК, в памяти которого был записан наилучший по тем временам набор
описаний изготовления минимального набора ферментов для синтеза белков. Основным
достоинством этих клеток было то, что они могли делиться и продолжать
накапливать информацию для изготовления все большего количества различных
белков, в результате чего в клетках появились органоиды. С этого момента
появились законченные растительные клетки. Пищеварения в те времена еще не было
и питание происходило только за счет избирательной трансмембранной диффузии
низко-молекулярных соединений из окружающей среды с последующим внутриклеточным
синтезом высокомолекулярных соединений.
Мы не будем далее развивать идеи возникновения клеточной жизни на Земле,
потому что с нас достаточно понимания, что первыми живыми существами были
одноклеточные растения, то, что мы называем системами с соматическим и
вегетативным БУС. Мы можем предположить, что эти первые одноклеточные растения
были самыми примитивными автотрофами (хемо- и фототрофами), питающимися простыми
химическими соединениями, потому что на Земле кроме камней и очень малого
количества органических соединений тогда ничего не было. Но их хватило, чтобы на
Земле «завязалась» жизнь.
Далее пошел обычный для Вселенной процесс захвата: одни клетки стали
захватывать другие. Если было равенство сил и возможностей клеток, то захват
приводил к объединению клеток в многоклеточные не дифференцированные
образования. При неравенстве сил и возможностей клеток могли возникать различные
ситуации:
поглощение и эксплуатация, т.е., использование свойств
поглощенных клеток для своих нужд. При этом поглощенные клетки оставались в
живых потому, что у клетки-хозяина еще не было необходимых энзимов и органелл
для переваривания поглощенной клетки. Так возникали дифференцированные
многоклеточные образования. Таким же путем хлоропласты «поселились» внутри
растительных клеток.
поглощение и переваривание, т.е., использование поглощенного
клеточного материала для своих нужд, когда у клеток-хищников уже
появились необходимые энзимы и органеллы для переваривания поглощенных клеток.
На первых порах внутриклеточного переваривания (пищеварения) не было, потому
что для этого необходимы специализированные внутриклеточные органеллы с
определенным набором специфических для каждого вида пищи энзимов. У первых
клеток, вероятно, были только рибосомы для синтеза белков и РНК и,
возможно, первичные вакуоли – не ограниченные мембраной органоиды
(сегодня это ограниченные мембраной органоиды), для хранения питательных веществ
клетки, получаемых из внешней среды путем трансмембранной диффузии, пиноцитоза
или фагоцитоза.
Но захват определенных клеток путем фагоцитоза и включение их в состав своей
внутренней среды приводил к кооперации этих клеток с телом (клеткой) хозяином и
увеличению числа специализированных органелл. Хлоропласты поглощают свет и СО2
и выделяют углеводы, которые могут быть усвоены клеткой-хозяином, которая, в
свою очередь, дает «дом» и защиту тем же хлоропластам. Таким же образом
появились внутриклеточные митохондрии. Наличие отдельных ДНК в хлоропластах и
митохондриях указывает на то, что когда-то они были отдельными организмами типа
бактерий, а сегодня они является составной частью клеток. Следовательно, на
первых порах захват привел к увеличению числа внутриклеточных специализированных
органелл.
Для своей жизнедеятельности первичные растения не требовали много энергии,
потому что для обмена веществ они довольствовались относительно простыми
химическими веществами, синтезируя из них более сложные белки, жиры и сахара. Но
появление многих органелл усилило синтетические способности и дало
клетке-хозяину высокомолекулярное питание, для которого необходимо было
появление новых комплектов энзимов. В клетке стали появляться все новые и новые
органеллы, увеличивающие возможности выживания клетки. Но увеличение числа
органелл привело к увеличению зависимости клетки от нормальной работы этих
органелл, которые неизбежно разрушались по мере их эксплуатации. Появилась
необходимость в их восстановлении, которая стала удовлетворяться усложнением
генного механизма, который сначала был механизмом только репликации
самой клетки. Как только этот механизм «научился» считывать не сразу всю
информацию с ДНК, а только ее часть, касающуюся разрушенной органеллы, этот
механизм стал также и регенеративным.
Следует отметить еще одно важное обстоятельство. Известно, что
в ДНК есть описание всех белковых молекул, из которых состоят внутриклеточные
структуры. Но неизвестно, где находится описание самих структур,
которые строятся из этих молекул. Мы не будем строить предположения о месте
нахождения этого описания и для удобства изложения примем априори, что оно
находится в той же молекуле ДНК, хотя нет никаких доказательств этого и вообще,
возможно, это не так. Так же неизвестен и механизм, каким образом из
реплицированных белков строятся эти структуры.
Первые одноклеточные появились на Земле более чем 3.6 млрд лет назад, еще в
эоархейскую эру. От них пошли бактерии и различные одноклеточный водоросли.
Спустя 1.3 млрд лет их эволюции (2.3 млрд лет назад) в результате
жизнедеятельности архейских анаэробных микроорганизмов в атмосфере Земли
образовалось большое количество кислорода, который, однако, был ядовит для
большинства из этих организмов (кислородная катастрофа [60]). Этим
была заложена предпосылка появления в последующем мира животных, для активного
передвижения которых необходимо было много энергии, которая могла получаться
только путем сгорания органики в кислороде. Но животные появились только лишь 3
млрд лет спустя. Первые надежные признаки появления животных датируются временем
примерно 500-600 млн лет назад, в эдиакарии. Следовательно, почти 3 млрд лет на
Земле «царствовали» только растения, эволюция которых прошла множество стадий:
от одноклеточных до многоклеточных, от автотрофов до гетеротрофов.
В литературе можно встретить описание «животных» докембрийского периода
[18, 26], но, вероятно, здесь допущена ошибка. Это были не животные, а
растения. Растения имеют только простой БУС, неспособный управлять
целенаправленным движением в окружающей реальности, потому что не
осознавал ее. Это значит, что растения не могли активно и целенаправленно
перемещаться в пространстве, такое перемещение характерно только для животных с
их сложными БУС (см. далее). Они либо должны быть неподвижно закреплены в почве,
на дне водоема или на хозяине (эпифиты и паразиты), либо быть свободно
подвешенными в толще воды, либо активно, но хаотически и нецеленаправлено
двигаться. Это предъявляет особые требования к их способу обмена веществ и их
строению. Основными способами обмена веществ у архаичных растений является
автотрофия (синтез высокомолекулярных соединений из низкомолекулярных за счет
энергии света, фотоавтотрофия, или энергии химических реакций,
хемоавтотрофия), которая была единственным способом пропитания в тех
местообитаниях, где мало питательных веществ [26]. Автотрофия не
требует наличия внутренних органов и органов передвижения. Описанные в
литературе вендобионты отличались именно таким строением тела, которое позволяло
им эффективно обмениваться веществами через поверхность их тела: они имели очень
большое соотношение поверхность-объем, и неподвижно лежали на дне, ожидая
питательные вещества, которые приносились к ним током воды (например, “стеганое
одеяло” Зейлахера[26]). На это же указывает отсутствие повреждений
тела систем, связанные с деятельностью животных (нет следов зубов и прочих
атрибутов животных), потому что в это время животных с их органами жевания
(зубы) еще не было. Растения органов не имеют, поэтому не было и зубов, хотя,
вероятно, уже в те времена были растения хищники (гетеротрофы), питающиеся
другими растениями, например, фитопланктоном. Поэтому то, что называют «эдиакарской
фауной» ([14]), на самом деле было «эдиакарской флорой» и первые
животные появились не в эдиакарии, а в кембрии, менее 500 млн лет назад.
Вероятно, можно также допустить наличие у некоторых из достаточно
«продвинутых» растений специальных приспособлений для активного перемещения,
некоторое подобие сократительных элементов (мышц), которые меняли форму тела
растений или приводили в движение дополнительные элементы типа жгутиков
(жгутиковые), но эти активные движения требовали больше энергии, чем в случае
пассивного и неподвижного образа жизни растений. Для этого растения должны были
стать хищниками – поглощать и переваривать другие растения (переваривать их
белки, жиры и углеводы). Однако эти активные перемещения должны были быть
нецеленаправленными в смысле перемещения тела в заданную
определенную точку пространства, потому что у них еще не было инстинкта,
направляющего движения (регулирующего поведенческие реакции). Они не «видели»
окружающую реальность и не могли знать, куда двигаться. Цель этих хаотических
движений была простой – увеличить вероятность встречи с пищей, но
не догнать ее. Эти растения могли быть хищниками, поедающими
другие растения (растительную плоть), но от этого они не переставали быть
растениями. Ведь никто не отрицает, что росянка круглолистная является
растением, хотя она плотоядный хищник, поедающий животных. В те времена животных
еще не было и в поедании одних растений другими нет ничего удивительного.
Кишечнополостные, ныне относимые к царству животных, очень хорошо подходят на
роль этих развитых растений, способных двигаться. У полипов даже есть зачаточная
нервная система, но это всего лишь диффузно разбросанные нервные клетки и не
ясно, есть ли у них инстинкт. Если есть инстинкт, то полипы являются животными с
сложным соматическим БУС (см. ниже), способными к целенаправленным
движениям. Если нет инстинкта, то они являются высшей формой растений, вернее,
переходной формой между растениями и животными, способной только к
нецеленаправленным хаотическим движениям. Морская актиния хаотически
шевелит своими щупальцами и есть много шансов на то, что они натолкнутся на
пищу. Но ее движения скорее хаотические, чем целенаправленные. То, что она
прячется от легкого прикосновения еще не указывает на стереотип движения (см.
ниже), это может быть также обычная реакция, подобная реакции систем с простым
блоком управления.
Поэтому высшими растениями являются не цветковые или покрытосеменные, а
двигающиеся и имеющие подобия органов, потому что из этих высших растений
произошли животные, когда они приобрели инстинкт, позволивший им двигаться в
окружающей реальности целенаправленно.
До появления жизни на Земле были только относительно простые химические
соединения и на таком «голодном пайке» выжить могли только растения. Но
появившись, растения дали остальному живому миру не только кислород, но и пищу,
что и определило появление мира животных, дав возможность вырабатывать
достаточно энергии для движений животных.
Функциональные возможности систем с вегетативным БУС.
Растения, в отличие от систем мира минералов, могут восстанавливать свои
разрушенные структуры, используя для этого систему обмена веществ.
Это отличие превращает абсолютную зависимость от разрушающего
действия окружающей реальности в относительную, потому что системы
научились восстанавливать свои разрушенные структуры. Это отличие возникает
только благодаря системе обмена веществ.
Возможности сознания простого соматического и вегетативного БУС.
Основным инструментом управляющих функций соматического БУС являются
простые соматические рефлексы, подобно простому БУС систем мира
минералов Они основаны на использовании РВВ для восприятия внешнего воздействия,
ППС для передачи сигнала в БПР, ССЭИ, РРД и ООС.
Основным инструментом управляющих функций вегетативного БУС являются
простые вегетативные рефлексы у растений, подобно простому БУС систем
мира минералов, и сложные вегетативные рефлексы (вегетативные рефлексы) у
животных (см. далее). Они основаны на использовании РОВ для восприятия внешнего
воздействия, ППС для передачи сигнала в БПР, АП, РРД и ООС.
Осознание своего Я. Вегетативное Я ощущает свое Я и осознает внутреннюю
реальность системы. Поэтому у вегетативного Я есть подсознание, но
нет сознания, потому что, оно не осознает окружающую реальность.
У растений, как и у систем всех последующих уровней, есть два своих Я: БПР
системы (Эго системы) и вегетативное Эго.
БПР системы (соматическое Я) ощущает свое Я, потому что знает свои
цели-стремления, оно знает «что Я такое», но не осознает
внутренней реальности системы, потому что не знает «где Я есть», потому что не
знает своей внутренней реальности.
Вегетативное Я системы системы также ощущает свое Я, потому что знает
свои цели-стремления, оно знает «что Я такое», но оно также и осознает
внутреннюю реальность системы, знает только часть ответа на вопрос: «где Я
есть». Но не осознает окружающую реальность и поэтому не знает ответа на вторую
часть вопроса : «где Я есть». Поэтому у вегетативного Я системы
есть предсознание, но нет сознания.
Наличие двух Я у систем с простым БУС и системой управления обмена веществ
объясняется тем, что произошло простое объединение двух систем по типу симбиоза
с разделением функций. Одна система «отвечает» за физическое «противостояние»
окружающей реальности, а вторая система обеспечивает восстановление разрушенных
структурных единиц первой системы. Таким образом первая система «зарабатывает»
постоянство своей структуры, противостоя окружающей реальности, а вторая систем
– постоянную защиту от разрушающего воздействия окружающей реальности, если это
воздействие уже произошло.
У растений оба Я, возможно, совершенно равноправны, потому что у них
одинаковые БУС – оба простые с равными возможностями, но с разными
точками приложения. Которое из них главнее, покажет их дальнейшая эволюция. Но у
растений это два относительно независимых Я и каждое из них занимается своим
делом, дополняя друг друга, хотя и не осознавая друг друга. Растения не «бегают»
и у них нет внутренних органов. Поэтому и у них нет такого вегетативного Я,
которое могло бы вмешиваться в деятельность соматического Я. Поэтому у растений
нет сна, он им не нужен (см далее).
Осознание внутренней реальности. Так как существуют два Я – соматическое
(БПР простого БУС) и вегетативное Я, необходимо рассматривать осознание
внутренней реальности для каждого из них в отдельности:
у соматического Я, подобно простому БУС систем мира минералов, нет
осознания внутренней реальности, потому что у него нет рецепторов, дающих
информацию о ней, и нет БД, в которой описана должная внутренняя реальность.
Поэтому он не может оценить ее и, следовательно, осознать,
у вегетативного Я есть рецепторы, дающие информацию о состоянии элементов
внутренней реальности, и у него есть БД, в которой описано должное состояние
элементов внутренней реальности, поэтому есть осознание своей
внутренней реальности, потому что есть ее оценка – есть предсознание.
Поэтому будем говорить только о вегетативном Я, поскольку соматическое Я
полностью подобно простому БУС у систем мира минералов и это описано выше. При
этом отметим, что есть вегетативные Я различных уровней развития: от простейшего
внутриклеточного у растений, до сложнейшего с вегетативной подкоркой головного
мозга и вегетативным инстинктом (см. далее). Многое, что будет написано ниже,
касается сложнейшего вегетативного Я человека.
Вегетативное Я уже не просто знает, что есть внутренняя реальность
системы – элементы исполнения системы, но уже знает, какой должна
быть эта внутренняя реальность. Оно может ее оценить, сравнив ее
состояние с тем, что должно быть (с «базой данных нутренней реальности», с ДНК),
а оценка – это осознание своей внутренней реальности. Осознание позволяет
определить, какие структуры разрушены и не могут выполнять свои функции, и
принять решение о действиях системы обмена веществ для их регенерации,
задействовав генный механизм восстановления. У этого Я есть предсознание.
Но это Я не знает окружающей реальности, поэтому у него нет сознания. Деревья
погружены во внутреннее созерцание и не замечают окружающей реальности.
Для того, чтобы восстанавливать разрушенные элементы системы БУС управляет
системой обмена веществ. Работа системы обмена веществ складывается из двух
составляющих:
координированная деятельность собственных элементов системы обмена веществ
и
сам обмен веществ, который складывается из многих процессов.
Вещества должны «приходить» в систему и «уходить» от нее, а внутри нее они
должны преобразовываться. Сама система практически ничего не делает для того,
чтобы вещества «приходили» к ней или «уходили» от нее. Она делает только
действия преобразования веществ в ней. Растения неподвижно стоят в почве или
свободно подвешены в толще воды и вещества поступают в них только путем притока
в растворенном состоянии. Растения могут и перемещаться током воды, но сами они
не передвигаются в сторону пищи (пассивное перемещение).
Вегетативное Я растений, в отличие от систем минерального мира, ощущает свою
внутреннюю реальность и полностью осознает ее. Оно ощущает свои внутренние
структуры, «знает», какими они должны быть и «знает», как восстанавливать эту
внутреннюю реальность с помощью обмена веществ, если они разрушены. У него нет
осознания строения самой системы обмена веществ, особенно в ее деталях, типа
генов и прочих ее атрибутов, но оно знает, когда и как ее задействовать, чтобы
регенерировать свои утерянные СФЕ или реплицировать самих себя в размножении.
Это уже предсознание, потому что есть ощущение (восприятие)
внутренней реальности и ее оценка – соотношение с самим
собой, т.е., определение, насколько внутренняя реальность отличается от должной.
Эти знания записаны в ДНК.
Это отличие превращает абсолютную зависимость от разрушающего
действия окружающей реальности в относительную, потому что системы
научились восстанавливать свои разрушенные структуры. Это отличие возникает
только благодаря системе обмена веществ.
Возможно кому-то покажется странным утверждение, что растения осознают
свою внутреннюю реальность. Но управление обменом веществ должно быть только
целенаправленным, потому что иначе растения не могли бы сохранять свою форму и
размеры. Целенаправленное управление обменом веществ возможно только при его
осознании. Если не принять осознание растениями внутренней реальности, то нужно
предположить, что кто-то или что-то внешнее следит за состоянием внутренней
реальности растений, определяет их повреждения и целенаправленно включает
необходимые элементы для регенерации повреждений. Кто-то или что-то должно это
делать, потому что само собой ничего не происходит. Или Бог (дух, жизненная сила
и пр.), или само растение, третьего не дано. Проще предположить, что само
растение занимается этим, чем предполагать вмешательство неизвестных сил. Если
определенное растение всегда имеет определенную форму, значит оно осознает то,
что оно делает, совершая процессы регенерации.
Осознания окружающей реальности нет, потому что для этого не хватает
ощущения и восприятия окружающей реальности и ее соотношения с собой.
Системы с такими БУС также являются объектами, потому что
однотипные системы всегда в ответ на одно и то же внешнее
воздействие дают одну и ту же реакцию, всегда делают одни и те же
действия, производят один и тот же результат действия. Их Я
абсолютно похожи, они всегда «хотят» одно и то же. Поэтому они объекты.
Наличие чувств и эмоций. У него нет чувств и эмоций, потому что нет
сознания.
Недостатки простого соматического и простого вегетативного БУС.
Их по прежнему три: шумовой дребезг ООС, зависимость от ситуации окружающей
реальности, потому что нет восприятия никакой информации о ней, и полуактивные
методы защиты от влияния окружающей реальности.
1. Шумовой дребезг ООС у систем мира растений имеет ту же
природу, что и у систем минерального мира – особенности ООС (квантованность
результата действия системы). Поэтому нет растений «каменного» типа (жестко
сохраняющих свою форму) и есть очень большое разнообразие в размерах и форме
растений. Сажаешь картофель и никогда заранее не знаешь, каким он вырастет.
2. Зависимость от ситуации окружающей реальности. Тем же
существенным недостатком систем с простым БУС является то, что они совершенно
ничего не «знают» об окружающей ситуации. Животное подойдет к растению и съест
его, огонь подползет и сожжет его, а оно не сделает ничего, чтобы спасти себя. И
это потому, что у систем мира растений нет:
информации об окружающей ситуации и они не знают, что чем
ситуация им угрожает (нет прогноза),
сознания, чтобы оценить эту информацию и сделать прогноз, хотя уже
есть обмен веществ, чтобы восстановить разрушенные структуры и
органов движения, чтобы иметь возможность изменить ситуацию
на основе прогноза, например, убежать или догнать (нет поведенческих
реакций).
3. Полуактивные методы защиты от влияния окружающей реальности.
Полуактивные в том понимании, что восстановление всегда возникает только после
появления разрушения внутренних структур. Система пассивно ждет и только после
внешнего воздействия активно восстанавливает разрушенные структуры. Это подобно
постоянному подкачиванию воздуха в проколотый баллон, вместо того, чтобы закрыть
дырку в баллоне.